Матричная цепь
Cтраница 3
В конце концов оказалось, что так оно и есть. Для синтеза фрагментов Оказаки в качестве затравок требуются, как выяснилось, короткие отрезки РНК, комплементарные матричной цепи ДНК. Эта РНК образуется в направлении 5 - 3 из ATP, GTP, СТР и UTP с помощью фермента, называемого примазой. К З - концу этой короткой одноцепочечной РНК-затравки и присоединяются друг за другом дезоксирибонуклеотидные остатки, комплементарные цепи-матрице ДНК. [31]
Цветные прямоугольники различной яркости и насыщенности, занимающие пятую и шестую полосы, предназначены для оценки цветопередачи на различных уровнях яркости, для контроля основных цветов кинескопа, а также для проверки коррекции предыскажений. Цветные прямоугольники, занимающие полосы 14 и 15, обладают полной насыщенностью и служат для оценки правильности работы матричных цепей в приемнике. [32]

Тиминовые димеры вырезаются, и брешь заделывается в результате последовательного участия четырех ферментов ( рис. 30 - 2), Первый из них, называемый УФ-эндонуклеазой, расщепляет поврежденную цепь с 5 -стороны от тиминового димера. Второй фермент - ДНК-полимераза I-достраивает к открытому З - концу этой цепи соответствующие дезоксирибонуклеотиды, образуя короткий отрезок ДНК, комплементарный матричной цепи. В ходе этого синтеза цепь, содержащая тиминовый димер, вытесняется. На третьей стадии эн-донуклеаза вырезает дефектный участок. [34]
 |
Возможная схема репликации генома вируса полиомиелита. [35] |
Наконец, необходимо указать, что РНК-геномы вируса полиомиелита и других пикорнавирусов способны участвовать в межмолекулярной рекомбинации. Скорее всего такая рекомбинация происходит по модели смены матриц: например, З - конец недостроенной ( -) нити отделяется от матричной цепи и перемещается на гомологичное место другой оказавшейся по соседству молекулы вирусной РНК. После завершения синтеза ( -) нити на новой матрице она может давать начало рекомбинантным ( - Ь) геномам. [36]
 |
Возможная схема репликации генома вируса полиомиелита. [37] |
Наконец, необходимо указать, что РНК-геномы вируса полиомиелита и других пикорнавирусов способны участвовать в межмолекулярной рекомбинации. Скорее всего такая рекомбинация происходит по модели смены матриц: например, З - конец недостроенной ( -) нити отделяется от матричной цепи и перемещается на гомологичное место другой оказавшейся по соседству молекулы вирусной РНК. После завершения синтеза ( -) нити на новой матрице она может давать начало рекомбинантным () геномам. [38]
После этого он катализирует отщепление пирофосфата, присоединяя одновременно новый нуклеотид к З - концу затравочной цепи. Работая таким образом, фермент может превратить одноцепочечную ДНК-матрицу в двухцепочечную ДНК, причем в каждой точке новосинтезированной цепи этой ДНК будет содержаться основание, комплементарное основанию матричной цепи. [39]
Транзистор - ы TW1, Т102 охвачены отрицательной обратной связью через резисторы R109, R209 и имеют пятикратное усиление. С движков линейных потенциометров R111 / R211, включенных как регулятор стереобаланса ( влево-вправо) сигналы подаются на базы транзисторов TW2, Т202, работающих как фазоинверторы для питания матричной цепи. [40]
ОН-группа концевого рибонуклеотида этой короткой цепи РНК служит затравкой для синтеза ДНК под действием ДНК-поли-меразы III. Точность синтеза определяется тем, что фермент редактирует синтезированную цепь: если ДНК-полимераза встраивает неправильный нуклеотид, то фермент сам может распознать неспособность этого нуклеотида образовать правильную пару с соответствующим нуклеотидом матричной цепи. Поскольку цепи антипараллельны, то на первой рост цепи совпадает с направлением движения репликативной вилки, а на второй - идет в противоположном направлении. [41]
Только одна из двух цепей ДНК подвергается транскрипции в ходе РНК-синтеза. Эту цепь называют темплатной цепью. Вторая комплементарная цепь ДНК называется матричной цепью. Поскольку цепь n - РНК формируется на темплатной цепи ДНК, эти цепи оказываются, в конце концов, комплементарными. Соответствующие отношения между цепями ДНК и РНК иллюстрируются ниже. [42]
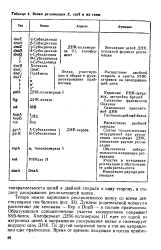 |
Белки репликации Е. colt и их гены. [43] |
Дуплекс родительской молекулы расплетают две хеликазы - Rep и DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК. По другой матричной цепи в том же направлении едет праймосома. [44]
 |
Белки репликации Е. colt и их гены. [45] |
Страницы: 1 2 3 4