Матричная цепь
Cтраница 4
Дуплекс родительской молекулы расплетают две хеликазы - Rep и DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК - По другой матричной цепи в том же направлении едет праймосома. [46]
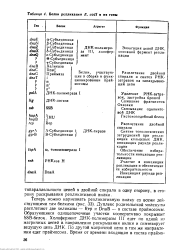 |
Белки репликации Е. cotl и их гены. [47] |
Дуплекс родительской молекулы расплетают две хеликазы - Rep к DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК. По другой матричной цепи в том же направлении едет праймосома. [48]
Наиболее хорошо изученной формой РНК являются небольшие 45-молекулы тРНК с мол. Похоже, что по размерам и основным структурным характеристикам тРНК бактериальных и эука-риотических клеток не различаются. Наличие адапторов, необходимых для переноса аминокислот в соответствующие участки матричной цепи мРНК, было предсказано еще до открытия тРНК - Предполагалось, что в молекуле адаптера должна иметься нуклеотидная последовательность, образующая антнкодон, который должен точно располагаться против соответствующего кодона, в каком-то связывающем участке белоксинтезирующей системы. Как теперь известно, молекулы тРНК действительно обладают предсказанными свойствами, однако при изучении их химических свойств было обнаружено много неожиданного. [49]
Дуплекс родительской молекулы расплетают две хеликазы - Rep и DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК - По другой матричной цепи в том же направлении едет праймосома. [50]
Для удаления ошибок репликации, неизбежных в процессе матричного синтеза таких огромных биополимеров, какими являются ДНК, существует специальная система ферментов репарации. Например, сопутствующие репликации одноцепочечные разрывы восстанавливаются при помощи ДНК-поли-меразы I и ДНК-лигазы. ДНК-полимераза I, будучи 3 - 5 -экзонуклеазой, проверяет правильность присоединения нуклеотидов вновь образованной нити ДНК к нуклеотидам матрицы и гидролизует концевой нуклеотид, если его основание не комплементарно основанию матричной цепи. ДНК-полимераза III, также обладающая нуклеазной активностью, будет добавлять нуклеотиды только в том случае, если предыдущее основание дочерней цепи комплементарно связано с соответствующим основанием матричной цепи. Таким образом, осуществляется репарация неправильного спаривания нуклеотидов и контролируется корректность синтеза ДНК. Наиболее полно изучены повреждения, возникающие в клетках под действием ультрафиолетового облучения. Оно вызывает, в частности, взаимодействие двух соседних пиримидиновых оснований, чаще всего тиминов. [51]
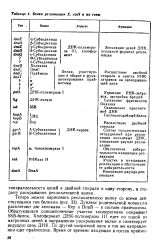 |
Белки репликации Е. colt и их гены. [52] |
Дуплекс родительской молекулы расплетают две хеликазы - Rep и DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК. По другой матричной цепи в том же направлении едет праймосома. [53]
Дуплекс родительской молекулы расплетают две хеликазы - Rep к DnaB - в составе праймосомы. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок. Холофермент ДНК-полимеразы III едет по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК. По другой матричной цепи в том же направлении едет праймосома. [54]
Репликация двухцепочечной ДНК, как известно, может осуществляться лишь в том случае, когда в полинуклеотидной цепи есть одиночный разрыв. При этом открывшаяся З - гидроксильная группа выполняет функцию затравки, и новая растущая цепь вытесняет старую комплементарную цепь, начиная с 5 -фосфатного конца. В одной из точек фермент может гойти с первоначальной матричной цепи и начать копирование комплементарной цепи. Это и влечет за собой образование разветвленной структуры. [55]
Для удаления ошибок репликации, неизбежных в процессе матричного синтеза таких огромных биополимеров, какими являются ДНК, существует специальная система ферментов репарации. Например, сопутствующие репликации одноцепочечные разрывы восстанавливаются при помощи ДНК-поли-меразы I и ДНК-лигазы. ДНК-полимераза I, будучи 3 - 5 -экзонуклеазой, проверяет правильность присоединения нуклеотидов вновь образованной нити ДНК к нуклеотидам матрицы и гидролизует концевой нуклеотид, если его основание не комплементарно основанию матричной цепи. ДНК-полимераза III, также обладающая нуклеазной активностью, будет добавлять нуклеотиды только в том случае, если предыдущее основание дочерней цепи комплементарно связано с соответствующим основанием матричной цепи. Таким образом, осуществляется репарация неправильного спаривания нуклеотидов и контролируется корректность синтеза ДНК. Наиболее полно изучены повреждения, возникающие в клетках под действием ультрафиолетового облучения. Оно вызывает, в частности, взаимодействие двух соседних пиримидиновых оснований, чаще всего тиминов. [56]
Большинство исследователей, занимающиеся биохимической генетикой и функциями нуклеиновых кислот, считали, что ДНК образуется только за счет репликации других молекул ДНК - Если транскрибирование РНК с ДНК может протекать свободно, то обратный процесс, а именно образование ДНК на РНК-матрице, считался маловероятным. Этот фермент обнаруживается в зрелых вирусных частицах. Наиболее тщательно очищенный фермент вирусов миелобластоза птиц состоит из двух белковых субъединиц, имеющих мол. Для функционирования фермента необходима короткая затравка и матричная цепь РНК - При этом сначала получается гибрид ДНК - РНК, из которого затем ( вероятно, после гидролитического расщепления цепи РНК под действием РНКазы Н, разд. [57]
Каким образом клеткам удается достичь столь высокой степени точности в выборе нужного основания в процессах репликации и транскрипции, а также при спаривании кодона с антикодоном в процессе синтеза белка. В ранних работах исследователи часто высказывали мнение, что специфичность спаривания оснований определяется исключительно образованием двух ( или соответственно трех) водородных связей и стабилизацией за счет взаимодействия соседних участков спирали. Оказалось, однако, что свободная энергия образования пар оснований мала ( гл. Исходя из современных энзимологических данных, можно предположить, что важную роль в обеспечении правильности спаривания играет сам фермент. Следовательно, связывающее место фермента может полностью окружить двойную спираль. На приведенном рисунке изображено гуаниновое основание матричной цепи молекулы ДНК, расположенное в месте наращивания комплементарной цепи ( ДНК или РНК) с З - конца. Для образования правильной пары оснований соответствующий нуклеозидтрифосфат должен быть пристроен до того, как произойдет реакция замещения, в результате которой нуклеотид присоединится к растущей цепи. Предположим, что у фермента есть связывающие места для дезоксирибозного компонента матричного нуклеотида и для сахарного компонента включающегося нуклеозидтрифоофата, причем эти места расположены на строго определенном расстоянии друг от друга. [58]

РНК-полимераза требует для своего функционирования присутствия ДНК. Фермент наиболее активен, если в качестве матрицы используется природная двухцепочечная ДНК из того же самого или из какого-либо другого организма. Из двух цепей ДНК транскрибируется только одна. В места, соответствующие остаткам аденина ДНК-матрицы, в новосинтезированную РНК включаются остатки урацила ( U), поскольку аденин и урацил образуют комплементарную пару. В места же, соответствующие тими-новым остаткам ДНК-матрицы, включаются остатки аденина. Остатки гуанина и цитозина в ДНК-матрице определяют включение соответственно цитози-новых и гуаниновых остатков в цепь РНК. Анализ нуклеотидного состава и нуклеотиднои последовательности новообразованной РНК показал, что она обладает противоположной по отношению к матрице полярностью и ее последовательность комплементарна последовательности матричной цепи. [60]
Страницы: 1 2 3 4