Глюкопиранозный цикл
Cтраница 1
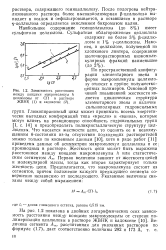 |
Зависимость расстояния. [1] |
Глюкопиранозный цикл может принимать восемь энергетически выгодных конформаций типа кресло и ванна, которые могут влиять на реакционную способность гидроксильных групп [ I, с. Что касается жесткости цепи, то оценить ее в невозмущенном 6-состоянии из-за отсутствия подходящих растворителей можно только по косвенным данным [10], хотя в ряде работ [11] приведены данные об асимметрии макромолекул целлюлозы и ее производных в растворе. Жесткость цепи может быть выражена расстоянием между концами макромолекулы h или статистическим сегментом Ат. Первая величина зависит от жесткости цепей и молекулярной массы, тогда как вторая характеризует только жесткость цепи. Предполагается, что макромолекула состоит из сегментов, причем положение каждого предыдущего сегмента совершенно независимо от последующего. [2]
Следовательно, макромолекулы ламинарина построены из d - глюкопиранозных циклов, которые соединены друг с другом ацетальной связью между 1 - м и 3 - м атомами углерода. Сильное отрицательное удельное вращение в щелочных растворах указывает на наличие в ламинарине р-связи глюкозных остатков. [3]
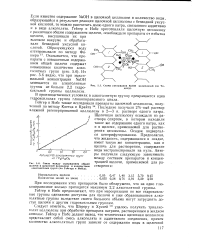 |
Схема связывания NaOH целлюлозой ( по Че-дину.| Связь между содержанием общей. [4] |
В производственных условиях в алкоголятную группу превращается одна гидроксильная группа глюкопиранозного цикла. [5]
Образование - ОСНОН - возможно, также вызывает дополнительные разрывы глюкопиранозных циклов, дальнейшую деполимеризацию молекулы целлюлозы и образование карбоксильных групп. Интенсивность этой полосы растет с увеличением дозы [2, 3, 95, 122, 154]: очевидно, она обусловлена карбоксильными группами. [6]
Окислительные реакции функциональных групп целлюлозы протекают по разным направлениям: окисление первичных спиртовых групп до альдегидных или карбоксильных, окисление вторичных спиртовых групп с разрывом или без разрыва глюкопиранозного цикла. Легче всего окисляются альдегидные группы редуцирующих концевых звеньев. Обычно окисление протекает одновременно по разным направлениям, и состав продуктов окисления весьма неоднороден. [7]
Все элементарные ячейки ( табл. 1.12) содержат два кристаллографически независимых ангидроцеллобиозных звена; связь ( 1 - - 4) между элементарными звеньями в каждой цепи приводит к чередованию глюкопиранозных циклов. В этом случае полярность центральной и угловых макромолекулярных цепей, которая может быть одинаковой или противоположной, не сказывается на полной симметрии. [8]
Необходимо отметить, что введение в молекулу целлюлозы одной и той же функциональной группы различными методами приводит к получению производных целлюлозы, в которых реакционная способность введен - % ых функциональных групп, в зависимости от положения их в молекуле ( в глюкопиранозном цикле или в боковой цепи), различна и соответственно отличаются и свойства получаемых продуктов. Это обстоятельство необходимо учитывать при выборе метода введения тех или иных функциональных групп. Систематическое изучение влияния положения функциональной группы в молекуле производного целлюлозы на химические, физико-химические и механические свойства этих производных представляет значительный интерес. [9]
При действии самой низкой дозы ( 0 52 - 1019 эВ / г) растворимость облученной целлюлозы со средневязкостной степенью полимеризации 3100 в 1 % - ном растворе едкого натра значительно увеличивается. Высокая растворимость облученной целлюлозы в 1 % - ном растворе едкого натра, вероятно, указывает на расщепление глюкопиранозных циклов, приводящее к дополнительному снижению молекулярного веса облученной целлюлозы. [10]
В конформации кресла 1C аксиальные и экваториальные заместители меняются местами. Экваториальные гидроксильные группы легче образуют гликозидные связи, легче этерифицируются и алкилируются, чем аксиальные. Конформация глюкопиранозного цикла достаточно жесткая и не меняется после введения объемистых заместителей. [11]
В то же время как в ( З - глюкопиранозе в конформации С1 все гидроксильные группы экваториальны, в а-глюкопиранозе в этой кон формации гидроксил у первого углеродного атома аксиален. Построение моделей показывает, что возникновение гликозидной связи между двумя остатками моносахарида в положении 1 - - 4 возможно только в том случае, когда оба гидроксила, участвующие в ее образовании, экваториальны. Поэтому построение а-глюкозидной связи между ангидридами глюкопиранозы в конформации С1 крайне затруднительно и мало вероятно. Ривз для глюкопиранозного цикла амилозы предложил конформации ванны 3В и В1, которые находятся в равновесии. [12]
В то время как в р-глюкопиранозе в конформации С1 все гидр-оксильные группы экваториальны, в а-глюкопиранозе в этой конформации гидроксил у первого углеродного атома аксиален. Построение моделей показывает, что возникновение гликозидной связи между двумя остатками моносахарида в положении 1 4 возможно, только в том случае, когда оба гидроксила, участвующие в ее образовании, экваториальны. Поэтому построение а-глюкозидной связи между ангидридами глюкопиранозы в конформации С1 крайне затруднительно и мало вероятно. Ривсом для глюкопиранозного цикла амилозы предложены конформации ванны 3В и В1, которые находятся в равновесии. В конформации 3В все гидроксилы, в том числе и глюкозидный, экваториальны, в конформации В 1 гидроксил у второго углеродного атома аксиален. [13]
В то же время как в 3-глюкопиранозе в конформации С1 все гидроксильные группы экваториальны, в ос-глюкопиранозе в этой кон-формации гидроксил у первого углеродного атома аксиален. Построение моделей показывает, что возникновение гликозидной связи между двумя остатками моносахарида в положении 1 - 4 возможно только в том случае, когда оба гидроксила, участвующие в ее образовании, экваториальны. Поэтому построение а-глюкозидной связи между ангидридами глюкопиранозы в конформации С1 крайне затруднительно и мало вероятно. Ривз для глюкопиранозного цикла амилозы предложил конформации ванны 3В и В1, которые находятся в равновесии. [14]

Страницы: 1 2