Элонгационный цикл
Cтраница 1
Элонгационный цикл начинается с поступления в рибосому Аа - тРНК при наличии там инициаторной ( формил) ме-тионил - тРНКр ( сразу после вышеописанной инициации) или пептидил-т РНК ( если рассматривать любой промежут. Предварительно вне рибосомы Аа - тРНК взаимод. ЕР-ТЦ) у прокариот или еЕР - 1 у эукариот; для взаимод. [1]
Идия элонгационного цикла оказывается блокированной. [2]
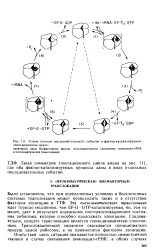 |
Схема полной последовательности событий в фактор-катализируемом элонгационном цикле. [3] |
Такая симметрия элонгационного цикла видна на рис. 111, где оба фактор-катализируемых процесса даны в виде отдельных последовательных событий. [4]
Хлорамфеникол ингибирует как естественную транспептидацию между пептидил-т РНК и аминоацил-т РНК в ходе элонгационного цикла, так и реакцию пептидил-т РНК или ее аналогов с пуроми-цином. Простейшее объяснение состоит в том, что хлорамфеникол является неактивным аналогом акцепторного субстрата и, связываясь с пептидилтрансферазным центром, конкурентно мешает взаимодействию настоящих акцепторов. Однако имеются указания также и на неконкурентный способ ингибирующего действия хлорамфеникола; и не исключено, что, связываясь, антибиотик подавляет какую-то катали. [5]
Известен целый ряд специфических ингибиторов, главным образом антибиотиков, блокирующих ту или иную стадию элонгационного цикла рибосомы. Большая часть из них действует избирательно либо на ррокариотические ( бактериальные), либо на эукариотические рибосомы. В соответствии с этим место специфического связьюания тетрациклина с рибосомой обнаружено на 30S субчастице, хотя при более высоких концентрациях он может связываться также и с 50S субчастицей, обнаруживая побочные действия. Интересно, что когда тройственный комплекс Aa - tRNA EF-TU GTP взаимодействует с 70S рибосомой в присутствии тетрациклина, то ГТФ Гидролизуется и EF-TU GDP освобождается, но аминоацил-т РНК не остается связанной. [6]
Все это делает невозможным даже приблизительную оценку изменения свободной энергии в самой реакции транспептидации в процессе нормального элонгационного цикла на рибосоме. Конечно, из того факта, что реакция протекает, и быстро, следует вывод об уменьшении свободной энергии рибосомного комплекса при транспептидации. Однако это уменьшение не может быть столь значительным, как - 30 кДж / моль ( - 7 ккал / моль) в стандартных условиях, а должно быть меньше по следующим соображениям. Во-первых, свободная энергия гидролиза связанного субстрата, при условии освобождения продуктов гидролиза, должна быть меньше таковой свободного субстрата, так как часть энергии должна была освободиться при связывании, если оно было спонтанным. Во-вторых, свободная энергия гидролиза субстрата, когда продукт задерживается в связанном виде, тоже должна быть меньше таковой в случае, когда продукт освобождается, так как в связанном состоянии продукта аккумулирована часть свободной энергии, если освобождение продукта является в принципе термодинамически спонтанным. Можно считать, что свободная энергия около - 30 кДж / моль ( - 7 ккал / моль), которая освободилась бы в реакции транспептидации в случае, когда субстраты и продукты свободны, здесь распределяется на предыдущую стадию связывания аминоацил-т РНК и последующую стадию транслокации, таким образом питая весь элонгационный цикл. [7]
Бесфакторное ( неэнзиматическое) связывание аминоацил-т РНК, катализируемая рибосомой транспептидация и бесфакторная ( неэнзи-матическая) транслокация создают бесфакторный элонгационный цикл. Повторение циклов дает в результате медленную бесфакторную элонгацию. В бесклеточных системах была проведена бесфакторная трансляция полиуридиловой кислоты и ряда синтетических гетеро-полинуклеотидов и показано полное соответствие полипептидного продукта матричному полинуклеотиду, согласно генетическому коду. [8]
Попеременно меняя концентрацию Mg2 в бесклеточной системе, можно было бы в известной мере имитировать действие факторов элонгации и тем самым значительно ускорить бесфакторный элонгационный цикл. [9]
Хотя наличие антикодон-кодонового сродства необходимо для лож - ого связьшания аминоацил-т РНК на рибосоме, это не значит, что частота ( ржного связывания будет пропорциональна константе сродства. С одной стороны, если в элонгационном цикле вслед зарбратимым Лдонзависимым связыванием тРНК быстро следует необратимая или едленно обратимая фаза ( например, запирание аминоацил-т РНК в It-участке или транспептидация), то ее скорость будет влиять на уровень ( Южного кодирования: чем выше скорость последующей необратимой [ медленно обратимой) фазы, тем выше ложное кодирование. Но если скорость следующей фазы цикла чересчур быстра и Сравнима со скоростью распада неправильного комплекса, то с большей вероятностью неправильная тРНК окажется вовлеченной в дальнейший икл. [10]
Это наводит на мысль о прямом действии вирусного компонента на какое-то звено элонгационного цикла. [11]
Вопрос об энергетике транслокации долгое время был еильно запутан ввиду ряда исторических причин и из-за традиционности мышления биохимиков. Дело в том, что участие ГТФ в процессе транслокации стало известно раньше всех прочих фактов, касающихся этой стадии элонгационного цикла. [12]
Очевидно, что любая энергия может затрачиваться либо для совершения полезной работы против термодинамического потенциала ( проведение процесса в гору), либо на преодоление барьеров в спонтанном ( с горы) процессе, без накопления полезной работы. Исключая первую альтернативу, остается признать, что вклад ГТФ является чисто кинетическим: сначала взаимодействие ГТФ с EF-G обеспечивает присоединение EF-G GTP к рибосоме и тем самым уменьшение барьеров в ходе транслокации, а затем гидролиз ГТФ снимает барьер, создаваемый самим EF-G для следующей стадии элонгационного цикла. Следовательно, энергия ГТФ тратится только на преодоление барьеров и в конечном счете целиком диссипирует в теплоту. Это и есть катализ транслокации. [13]
Естественно поставить вопрос, что служит ведущим актом - перемещение матрицы или перемещение пептидил-т РНК. Ряд фактов указывает на то, что ведущим является транслокационное перемещение тРНК, которая своим антикодоном тянет за собой кодон матрицы. В качестве одной из ярких демонстраций независимости транслокации от матричного полинуклеотида можно рассматривать открытие рибо-сомного синтеза полипептида из аминоацил-т РНК в отсутствие матричного полинуклеотида; в этом случае было показано существование нормального элонгационного цикла с EF-G GTP-катализи-руемой транслокацией. [14]
EF-TU ( так же как и эукариотический EF-1) способен взаимо - йствовать с аналогом, так что белок приобретает сродство к амино - ИИл-тРНК. Белок, аналог ГТФ и аминоацил-т РНК образуют тройствен - jfafi комплекс. Тройственный комплекс связывается с рибосомой. Однако 1едующая реакция ( 2) не происходит, EF-TU продолжает сохранять одство к аминоацил-т РНК и рибосоме, и аминоацил-т РНК остается 1деть в рибосоме вместе с белком и нерасщепляемым аналогом ГТФ. РНК неспособна служить акцепторным суб - ратом в реакции транспептидации с пептидил-т РНК и следующая 1адия элонгационного цикла оказьюается блокированной. [15]
Страницы: 1 2