Инициация - раунд - репликация
Cтраница 1
 |
Схема репликации генома фага Т7. [1] |
Инициация раунда репликации происходит внутри молекулы - на расстоянии, примерно соответствующем 15 % длины генома от одного из концов, условно называемого левым. Здесь имеются промоторы, которые узнаются фагоспецифической ДНК-зависимой РНК-полиме-разой. Без транскрипции этого оп-района раунд репликации не начинается. [2]
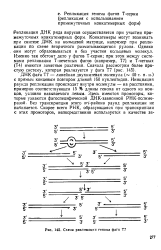 |
Схема репликации генома фага Т7. [3] |
Инициация раунда репликации происходит внутри молекулы - на расстоянии, примерно соответствующем 15 % длины генома от одного из концов, условно называемого левым. Здесь имеются промоторы, которые узнаются фагоспецифической ДНК-зависимой РНК-полиме-разой. Без транскрипции этого ori - района раунд репликации не начинается. [4]
 |
Схема репликации генома фага Т7. [5] |
Инициация раунда репликации происходит внутри молекулы - на расстоянии, примерно соответствующем 15 % длины генома от одного из концов, условно называемого левым. Здесь имеются промоторы, которые узнаются фагоспецифической ДНК-зависимой РНК-полиме-разой. Без транскрипции этого оп - района раунд репликации не начинается. [6]
 |
Структура реплицирующейся бактериальной хромосомы. [7] |
При высокой скорости роста бактерий инициация нового раунда репликации происходит еще до окончания предыдущего. [8]
 |
Структура реплицирующейся бактериальной хромосомы. [9] |
При высокой скорости роста бак тернй инициация нового раунда репликации происходит еще до окончания предыдущего. [10]
По крайней мере у бактерий топоизомеразы играют важную роль не только в ходе репликации и на завершающих стадиях этого процесса, но и при инициации раунда репликации. [11]
Другими словами, репликация идет без выщепления резидентного профага и, по существу, представляет собой реплика / пивную транспозицию. Выступающие 5 -концы клеточной ДНК соединяются с 3 -концами вирус-специфических последовательностей, а З - концы клеточной ДНК выполняют роль затравки. Таким образом, инициация раунда репликации представляет собой в этом случае вариант рекомбинационной инициации. В результате полуконсервативной репликации н последующих процессов репарации в клеточной хромосоме оказывается две копии профага; в каждой из них одна из цепей происходит из резидентного профага, а вторая синтезирована заново. При повторении этого процесса количество профагов в клеточной хромосоме может достигать сотни. [12]
Другими словами, репликация идет без выщепления резидентного профага и, по существу, представляет собой репликативную транспозицию. Выступающие 5 -концы клеточной ДНК соединяются с З - концамн вирус-специфических последовательностей, а З - концы клеточной ДНК выполняют роль затравки. Таким образом, инициация раунда репликации представляет собой в этом случае вариант рекомбинационной инициации. В результате Полуконсервативной репликации и последующих процессов репарации в клеточной хромосоме оказывается две копии профага; В каждой из них одна цз цепей происходит из резидентного профага, а вторая синтезирована заново. При повторении этого процесса количество профагов в клеточной хромосоме может достигать сотни. [13]
Что касается инициации на внутренних участках двухнитевой матрицы, то здесь также нужно различать два основных способа. Однако синтез затравки возможен только в том случае, если матрица соответствующим образом подготовлена. Подготовка включает взаимодействие между вирус-специфическими белками, регулирующими инициацию раунда репликации, и специфическими участками инициации репликации ori ( от англ, origin - начало) в молекуле ДНК - Например, с участком ori в ДНК фага h первично взаимодействует фагоспецифический белок - О, с белком О взаимодействует другой фагоспецифический полипептид - белок Р, который В свою очередь образует комплекс с одной из клеточных хеликаз - продуктом гена dna В. [14]
Что касается инициации на внутренних участках двухнитевой матрицы, то здесь также нужно различать два основных способа. Однако синтез затравки возможен только в том случае, если матрица соответствующим образом подготовлена. Подготовка включает взаимодействие между вирус-специфическими белками, регулирующими инициацию раунда репликации, и специфическими участками инициации репликации ori ( от англ, origin - начало) в молекуле ДНК - Например, с участком ori в ДНК фага X первично взаимодействует фагоспецифический белок - О, с белком О взаимодействует другой фагоспецифический полипептид - белок Р, который свою очередь образует комплекс с одной из клеточных хеликаз - продуктом гена ana В. [15]
Страницы: 1 2