Вращение - зонд
Cтраница 1
Вращение зонда в таких сетчатых системах при температурах ниже Тст практически изотропно, а зависимости lgrc от 1 / Т при различной концентрации ФГЭ ( с увеличением содержания ФГЭ густота сетки уменьшается) описываются аррениу-совскими прямыми с увеличивающимся наклоном. Рост Е с разряжением сетки указывает на то, что движение зонда сопровождается перестройкой все более крупных кинетических единиц. В пользу этого свидетельствует линейная зависимость - Igto от Е сетчатого полимера. Для меток в области температур ниже Тст характер зависимостей от концентрации ФГЭ такой же, как для зонда. Вблизи Тст зависимость lgrc от 1 / 7 меток претерпевает излом. Происходит изменение характера движения метки от мелкомасштабного к сегментальному с большей энергией активации, которая, наоборот, уменьшается по мере разрыхления сетки. Зонд слабо чувствителен к появлению сегментального движения, поскольку локализуется преимущественно в дефектных областях сетки ( дырках), размер которых значительно превышает объем радикала. Вместе с тем TC зонда растет, а Е падает с увеличением густоты сетки, что может свидетельствовать о росте структурной неоднородности системы: доля крупных дырок увеличивается при одновременном уменьшении общей доли свободного объема за счет более сшитых участков. [1]
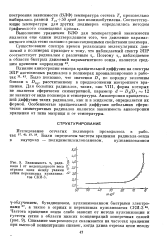 |
Зависимость тс ради-кала I от молекулярного веса отрезка цепи между узлами сетки перекисных вулканиза-тов СКИ-3. [2] |
Частота вращения зонда слабо зависит от метода вулканизации и густоты сетки в области низких концентраций поперечных связей ( рие. [3]
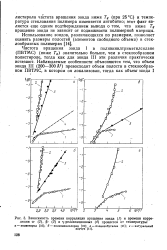 |
Зависимость времени корреляции вращения зонда ( 1 и времени корреляции а - ( 2, Р - ( 3 и - релаксационных ( 4 процессов от температуры. [4] |
Частота вращения зонда I в поливинилтриметилсилане ( ПВТМС) ( ниже Tg) значительно больше, чем в стеклообразном полистироле, тогда как для зонда III эти различия практически исчезают. [5]
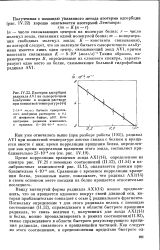 |
Изотерма адсорбции радикала AVI на сывороточном альбумине в водном растворе при комнатной температуре. [6] |
Время корреляции вращения зонда АХЦ14), определенное из спектра рис. IV. Сравнение с временем корреляции вращения белка, указанным выше, показывает, что радикал АХ1 ( 14) в своем основном месте посадки на белке вращается относительно последнего. [7]
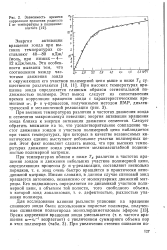 |
Зависимость времени. [8] |
Энергии активации вращения зонда при высоких температурах составляют 40 - 80 кДж / / моль, при низких - 4 - 12 кДж / моль. [9]
При высоких температурах вращение зонда определяется главным образом сегментальной подвижностью полимера. [10]
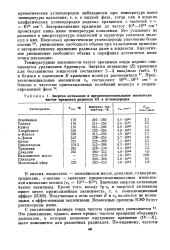 |
Энергия активации и предэкспоненциальные множители частот вращения радикала III в углеводородах. [11] |
Температурные зависимости частот вращения зонда хорошо описываются уравнением Аррениуса. [12]
Эффективная энергия активации вращения зонда уменьшается с увеличением концентрации поперечных связей. [13]
Зависимость времени корреляции вращения зонда АХЩ14) от температуры свидетельствует об изменении подвижности углеводородных цепей детергента. Сравнительно резкое уменьшение величины при переходе из геля в жидкокристаллическое состояние отражает состояние этих цепей при их плавлении. Отсутствие же резкого изменения величины т в области перехода II свидетельствует в свою очередь об отсутствии сколь-либо существенного изменения подвижности углеводородных цепей при переходе между жидкокристаллическими фазами. [14]
В пленках МЭА частота вращения зонда резко возрастает, а для пленок МТПА и МПА - меняется незначительно. [15]
Страницы: 1 2 3 4