Вращение - вокруг одинарная связь
Cтраница 3
Конечно, ценность такого рода оценок снижается вследствие того, что влияние растворителя в них никак не учитывается, и полученные цифры справедливы для вакуума. В водных растворах полинуклеотидов гидрофобные взаимодействия должны дополнительно стабилизировать укладку оснований в стопку. Действительно, стабилизации невалентными взаимодействиями ( 5 - 8 ккал / моль-основание) еще недостаточно, чтобы сделать энергию системы полинуклеотид - растворитель минимальной для этой конформации: в органических растворителях ( например, поли - С в этиленгликоле [36]), которые, как известно, являются эффективными денатуратами, минимум свободной энергии соответствует беспорядочной кокформацки нолинуклеотида. За возникновение беспорядочной конформации ответственна при этом конфигурационная энтропия, которая резко повышается при размораживании вращений вокруг одинарных связей рибозофосфатного скелета. [31]
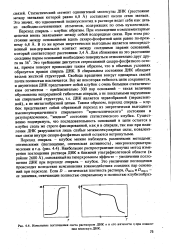 |
Изменение поглощения света раствором ДНК а и его вязкости т. при плавлении молекул ДНК. [32] |
При охлаждении комплементарные цепочки вновь завязывают между собой водородные связи. При этом расстояние между основаниями вдоль сахаро-фосфатной цепи равно по-прежнему 6 8 А. В то же время энергетически выгодным является непосредственный ван-дер-ваальсов контакт между соседними парами оснований, который соответствует расстоянию 3 4 А. В спиральном состоянии ДНК обладает весьма жесткой структурой. Свобода вращения вокруг одинарных связей почти полностью исключается. Все же некоторая гибкость цепи сохраняется, и двунитевая ДНК представляет собой клубок с очень большим статистическим сегментом - приблизительно 300 пар оснований - такая величина обусловлена непрерывной гибкостью спирали, а не локальными нарушениями спиральной структуры, т.е. ДНК является червеобразной ( персистент-ной), а не зигзагообразной цепью. Таким образом, переход спираль - клубок представляет собой обратимый переход из энергетически выгодного высокоупорядоченного спирального кристаллического состояния в разупорядоченное, жидкое состояние статистического клубка. Существенно подчеркнуть, что последовательность оснований в цепи остается в клубке столь же строго фиксированной, как и в спирали, так как при плавлении ДНК разрушаются лишь слабые межмолекулярные силы, ковалент-ные связи внутри сахаро-фосфатных цепей остаются нетронутыми. [33]
Это особенно важно в связи с теорией Гриффитса, согласно которой любая трещина или дефект в материале приводят к росту напряжения. Полимерные стекла менее хрупки, чем неорганические, потому что молекулы их имеют некоторую свободу движения и перегруппировки под действием приложенного напряжения, чего нет в неорганических стеклах. Последние почти совершенно упруги, у них отсутствует пластическая деформация, присущая металлам. Полимерные стекла ( и полимеры вообще) не так идеально упруги; им свойственна некоторая необратимая пластическая и вязкотекучая деформация, которая способствует рассеянию энергии упругой деформации. Необратимые деформации такого типа являются следствием существования вторичных, довольно слабых сил взаимодействия между молекулами, благодаря чему становится возможным вращение вокруг одинарных связей в молекулах при температурах ниже температуры стеклования. [34]
Страницы: 1 2 3