Ганглий
Cтраница 3
Белый ramus communicans состоит из предганглиевых аксонов: на пути их ко входу в симпатический ганглий. Имеется также серый ramus communicans, состоящий преимущественно из аксонов симпатических нейронов, которые должны исходить из висцеральной полости, чтобы пройти к потовым железам кожи, мускулам волос и кровеносных сосудов. [31]
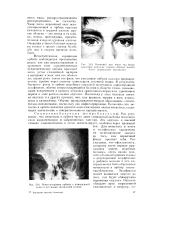 |
Нпетпний вид через год после.| Рентгенограмма орбиты с помещенной в нее 5 лет назад танталовой сеткой. [32] |
Риз установил, что метастазы в орбиту часто дают симпатикобластомы мозгового слоя надпочечников и забрюптипных ганглий. [33]
Исследованиями [435] также показано, что большие ганглии неправильной формы являются конгломератами нескольких более мелких ганглий более правильной формы. [34]
У позвоночных состоит из череппомозговых и спинномозговых нервов, а также расположенных по их ходу ганглиев. ЦНС с кожей, мышцами, сухожилиями, сенсорными органами, относятся к СНС, а нервы, связывающие ЦНС с внутр. ЦНС претерпевает значительно меньшие эволюц. [35]
Афферентные сигналы в гипоталамус поступают из коры больших полушарий, из таламических структур, ядер базальных ганглиев. Одним из основных эфферентных путей является медильный мозговой пучок, или паравентрикулярная система, и мамилло-тегментальный тракт. Волокна этих путей идут в каудальном направлении по стенкам водопровода мозга или сильвиева водопровода, дают многочисленные ответвления к структурам среднего мозга. Аксоны клеток гипоталамических ядер образуют также большое количество коротких эфферентных путей, идущих в таламическую и субтала мическую области и в другие подкорковые образования. [36]
Центральная нервная система у таракана менее компактна, чем у млекопитающих; она представляет собой ряды ганглиев. Эти ганглии не аналогичны периферическим ганглиям млекопитающих ( некоторые являются лишь местом расположения синапсов вегетативной нервной системы), скорее они соответствуют головному и спинному мозгу млекопитающих. Подобно им ганглии насекомых содержат координирующие системы и защищены от влияния многих посторонних веществ. [37]
При вытеснении нефти водой при движении без перемешивания большая доля нефти защемляется в виде нефтяных капель нефтяных ганглиев. Ганглий образуется на продвигающемся вперед фронте, когда часть нефти окружается водой и узкая шейка, соединяющая основное количество нефти с данным объемом, разрывается. [38]
Щетинкочелюстное Sagitta, схема строения: / - хитиноид-ные крючки-щетинки; 2 - мозг; 3 - брюшной ганглий; 4 - кишка; 5 - яичник; 6 - семенник; 7 - хвостовой плавник. [39]
Отравление организма приводит, как правило, к деформации отдельных органов и тканей - жирового тела, нервных ганглиев. Отмечается разрушение форменных элементов гемолимфы. При этом уменьшается количество пролейкоцитов и макронук-леоцитов и значительно увеличивается число патологических и мертвых клеток. Патологическая деформация нервной системы затрагивает также область нейросекреторных клеток, вследствие чего нарушается синтез и выделение нейросекрета. Выделяемый нейросекрет заметно теряет обычную в норме гранулярную структуру и превращается в гомогенизированную, интенсивно окрашивающуюся массу. [40]
Холинергическими являются и нейроны спинного мозга, иннервирующие хромаффин-ные клетки, а также преганглионарные нейроны, иннервирующие нервные клетки интра-муральных и экстрамуральных ганглиев. Полагают, что олинергические нейроны имеются в составе ретикулярной формации среднего мозга, мозжечка, базальных ганглиях и коре. [41]
Изменение структуры порового пространства при защемлении нефти определяется как порометрической харак7е - ристикой коллектора, так и распределением защемленных нефтяных ганглий по размерам. [42]
По-видимому, в механизме арминового спазма наряду с антихолин-эстеразным эффектом имеет значение и прямое холиномиметическое действие армина на холинорецепторы клеток ганглиев блуждающего нерва или самих гладких мышц бронхов. [43]
Таким образом, по предложенному механизму реализуются большие значения виброускорений в пласте, которые приводят к вовлечению остаточной нефти в форме ганглиев и глобул в фильтрационный поток. [44]
Частый ритм возбуждений преганглионарных волокон, превышающих частоту естественных импульсов, возникающих в нейронах вегетативной нервной системы, частично блокируется в синапсах ганглиев и постганглионарные волокна возбуждаются в более редком ритме. Таким образом, обнаруживается трансформация ритма нервных импульсов. Частота стимуляции свыше 100 в секунду вызывает полную блокаду проведения через синапс. [45]
Страницы: 1 2 3 4