Гипохромизм
Cтраница 1
Гипохромизм в применении к спектру димера предполагает, что полная сила осциллятора ( если оценить ее по площади спектральной полосы) перехода в димере меньше, чем удвоенная сила осциллятора соответствующего перехода в мономере; обратное явление называется гиперхромизмом. В рассмотренном случае интенсивности спектральных полос димера составляют 85 - 90 % интенсивностей соответствующих полос мономера. Таким образом, димер при переходе получает от мономера лишь часть силы осциллятора. Это означает, что в стабильность димера частично вносит вклад и энергия взаимодействия с высоко лежащими возбужденными состояниями. Поскольку симметрия этих состояний скорее всего отлична от симметрии низко расположенных состояний, общий эффект состоит в уменьшении силы осциллятора димера. Например, Фергюсон и др. [104] показали, что смешивание синглетного состояния В2и с энергией 44000 см 1 с состоянием 2ы, связанным с переходом вдоль короткой оси с энергией 26000 см 1, объясняет гипохромизм в спектре 9-хлорантрацена. Если резонансная энергия р ( определяемая формулой (1.4.2.06)) отрицательна, что обычно имеет место, то, поскольку наблюдается сдвиг в красную сторону ( как видно на рис. 1.4.6), атомы хлора находятся в / / троне-конфигурации. Таким образом, если атомы хлора находятся в ноконфигурации, связь в димере ослабляется. [1]
Гипохромизм важен не только сам по себе, как чрезвычайно интересное оптическое явление, но главным образом как феномен, дающий нам в руки простой и удобный метод, который можно использовать в химии нуклеиновых кислот для качественной и количественной оценки процессов ориентации - дезориентации ( таких, как денатурация, ренатурация, обратимое образование гомополимерных комплексов или образование гибридных спиралей ДНК - РНК), а также для установления генетической связи между ДНК из различных организмов или из различных клеток одного и того же организма. Все, что требуется для проведения такой оценки - это спектрофотометр или какой-нибудь другой прибор, с помощью которого можно измерять поглощение света в области 260 ммк. Первый максимум поглощения у всех исследованных видов ДНК располагается в интервале 256 - 265 ммк; вблизи 230 ммк находится минимум, а второй максимум поглощения лежит в далекой ультрафиолетовой области, при 195 ммк. [2]
Теория гипохромизма основывается на учете электростатического диполь-дипольного и поляризационного взаимодействий дипольных моментов перехода в упорядоченной системе биополимера. [3]
Гидролизом устраняется гипохромизм и нивелируется оптическая плотность НК, обусловленная структурным состоянием их молекул и особенно состоянием азотистых оснований. Часть белка остается в гадролизатах, поэтому содержание НК определяется по разности поглощения гадролизата при 270 ммк. [4]
Как известно, гипохромизм двойной спирали растет с длиной цепи. Однако разные типы РНК и денатурированной ДНК получены из разных источников и отличаются составом и последовательностью оснований и, по-видимому, распределение одноцепочечных и двухцепочечных участков в них различно. [5]
Даже в области Яшш гипохромизм претерпевает значительные количественные изменения. [6]
Очевидно, что исчезновение гипохромизма при переходе спираль - клубок может дать количественную меру а-спи-ральности белка. [7]
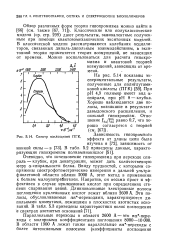 |
Спектр поглощения ПГК - Зависимость гипохромного. [8] |
Очевидно, что исчезновение гипохромизма при переходе спираль - клубок, при денатурации, может дать количественную меру а-спиральности белка. Ввиду трудностей, с которыми сопряжены спектрофотометрические измерения в дальней ультрафиолетовой области вблизи 2000 А, этот метод в применении к белкам малоупотребителен. Напротив, он весьма прост и эффективен в случае нуклеиновых кислот при определениях степени спаривания цепей. [9]
При увеличении длины цепи олигонуклеотида гипохромизм быстро достигает предельного значения. [10]
Упорядочение структуры молекул тРНК сопровождается появлением гипохромизма ( до 22 %) [64], увеличением числа медленно обменивающихся протонов [74], уменьшением числа оснований, доступных для формальдегида [64] и различных нуклеаз [75], увеличением коэффициента седиментации и уменьшением характеристической вязкости. [11]
Весьма вероятно, что основной причиной гипохромизма является прямое взаимодействие гетероциклических оснований и что влияние на этот эффект образования водородных связей обусловлено в первую очередь укладкой нуклеотидов в упорядоченную структуру с фактически полным ограничением движения по сравнению с оли-гонуклеотидами. В то время как при или вблизи А Макс оптическое поглощение понижается, при больших длинах волн ( приблизительно при 280 - 300 MI для производных аденина и урацила) оно незначительно возрастает. [12]
У полинуклеотидов и спиральных комплексов, кроме гипохромизма при Ямакс, возникающего в результате упаковки друг над другом л-л хромофорных единиц, гиперхромный эффект проявляется и при более длинных длинах волн. Такое увеличение оптического поглощения ( при 280 - 300 мц) замечено у нескольких спиральных полинуклеотидных комплексов. [13]
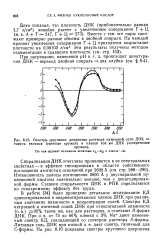 |
Спектры кругового дихроизма раствора натриевой соли ДНК из тимуса теленка ( светлые кружки и пленки той же ДНК ( зачерненные. [14] |
Спирализация ДНК отчетливо проявляется в ее спектральных свойствах - в эффекте гипохромизма в области собственного поглощения азотистых оснований при 2600 А ( см. стр. Интенсивность полосы поглощения 2600 А у двуспиральной нуклеиновой кислоты значительно меньше, чем у денатурированной формы. Степени спиральности ДНК и РНК определяются по гипохромному эффекту без труда. [15]
Страницы: 1 2 3