Гликогенолиз
Cтраница 3
Анаэробный распад гликогена ( гликогенолиз) или глюкозы ( гликолиз) начинается с этапа образования глюкозофосфорпых эфиров. При гликогенолизе вначале образуется глюкозе-1 - фосфат, который затем под влиянием фермента фосфоглюкомутазьг превращается в глю-козо-0 - фосфат. При фосфорилировании глюкозы процесс начинается с образования глюкозо - С-фосфата. [31]
Глюкагон является гормоном-антагонистом инсулина. Он стимулирует гликогенолиз и липолиз, а также активирует процесс глюконеогенеза. Глюкагон взаимодействует с клетками-мишенями по мембрано-опосредованному механизму ( гл. Через вторичный посредник - цАМФ он активирует протеинкиназу, киназу фосфорилазу и фосфорилазу Ь, что приводит к мобилизации глюкозы из гликогена. Как и инсулин, глюкагон регулирует метаболические процессы преимущественно в печени, мышцах и жировой ткани. [32]
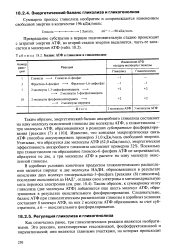 |
Баланс АТФ в гликолизе и гликогенолизе. [33] |
Известно, что концевая макроэргическая связь АТФ способна аккумулировать примерно 31 0 кДж / моль свободной энергии. Поскольку при гликогенолизе на образование глюкозо-6 - фосфата АТФ не затрачивается, образуется не две, а три молекулы АТФ в расчете на одну молекулу окисленной глюкозы. [34]
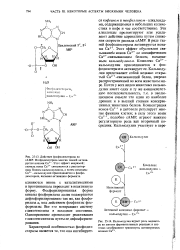
Фосфорилированная форма киназы фосфорилазы далее подвергается дефосфорилированию так же, как фосфо-рилаза а, под действием фосфатазы фосфорилазы. Все это возвращает систему гликогенолиза в исходное состояние. Одновременно происходит реактивация гликоген-синтазы путем ее дефосфорили-рования. [36]
 |
Аминокислотная последовательность глюкагона человека и быка ( верхняя и секретина свиньи ( нижняя. [37] |
О химии гормональных рецепторов известно мало; они, по-видимому, являются белками или конъюгатами белков. В случае глюкагона ( табл. 24.2.4), стимулирующего гликогенолиз в печени, имеются некоторые данные об аминокислотных остатках, ответственных за биологическую активность. Так, глюкагон, лишенный Af-концевого остатка гисти-дина, связывается с рецептором в печени, но не активирует аде-нилатциклазу. Фрагменты глюкагона, содержащие остатки 1 - 21 и 20 - 29, не связываются рецептором и не активируют фермент. His-1, по-видимому, важен для биологической активности. Интересно, что секретин ( см. табл. 24.2.4), также содержащий А - кон-цевой гистидин, и глюкагон стимулируют аденилатциклазную систему в жировых клетках крыс, но посредством различных рецепторов. Взаимодействие между глюкагоном и рецептором, по крайней мере частично, осуществляется посредством гидрофобных остатков близ С-конца гормона. Однако в отличие от других полипептидных гормонов типа р-кортикотропина и гастрина в случае глюкагона важна, по-видимому, вся структура. [38]
В задаче предлагается исследовать влияние добавления АДФ на интенсивность гликогенолиза. [39]
В результате всех этих работ выяснилось, что активный дрожжевой сок содержит не один фермент, а с л о ж ную ферментную систему, состоящую из большого числа компонентов как белковой, так и небелковой природы. Сейчас известно, что в осуществлении, например, гликогенолиза принимает участие более 10 отдельных ферментов и несколько термостабильных коферментов. Другими словами, то, что раньше называлось зимазой и понималось как один фермент, в действительности представляет целую систему ферментов брожения. [40]
Переход АТФ в АДФ связан с потерей АТФ одного остатка фосфорной кислоты. После образования глюкозо-6 - монофосфорного эфира дальнейшие пути гликолиза и гликогенолиза вполне совпадают и могут быть рассмотрены совместно. Из последнего под влиянием особого фермента фосфофруктокиназы путем перефосфорилирования с аденозинтрифосфорной кислотой возникает фруктоз о-1 6 - д и ф о с-ф о р н ы и эфир Л. А. Иванова ( IV), которому английские авторы несправедливо присвоили название эфира Гарден-Ионга. [41]
Рг-адрено-рецепторы, ответственные за расслабление неисчерченных мышц сосудов, бронхов, матки н гликогенолиз. Наконец, в последние годы получены убедительные доказательства существования пре - и постсннаптически расположенных ai - и а2 - адренорецепторов [ Langer S. [42]
Во время опасности или какого-либо стресса ( нервного напряжения) адреналин моментально выбрасывается в кровяное русло. Действуя, как полагают, в качестве активатора соответствующего фермента, он стимулирует гликогенолиз в печени. Освобождающаяся глюкоза появляется в кровяном русле в очень больших количествах, и содержание сахара в крови настолько повышается, что сахар начинает выделяться с мочой. Благодаря такому стремительному появлению глюкозы в крови организм получает основное питательное вещество именно в тот момент, когда оно ему крайне необходимо. [43]
Молекула киназы фосфорилазы состоит из субъединиц четырех типов: сфуб. Молекулярная масса фермента - 1 3 - 106 Да - отвечает формуле ( сфуб) - Киназа фосфорилазы играет, как показано, ключевую роль в регуляции обмена гликогена и в сопряжении гликогенолиза и мышечного сокращения. Активация киназы достигается фосфорилированием а - и р-субъединиц, которое катализирует цАМФ - зависимая протеинкиназа. Каталитическую роль выполняет у-субъединица; б-субъединица идентична Са2 - связывающему белку - кальмодулину. Ферментативная активность киназы фосфорилазы полностью зависит от ионов Са2 На р-субъединице фермента имеется регуляторный центр, обладающий высоким сродством к АДФ. [44]
Для ишемизированного миокарда характерны сниженное окислительное фосфорилирование и повышенный анаэробный обмен. Раннее увеличение гликогенолиза и гликолиза за счет имеющегося в сердечной мышце гликогена и глюкозы, усиленно поглощаемой миокардом в начальной стадии ишемии, происходит в результате повышения внутриклеточной концентрации катехоламинов и цАМФ, что в свою очередь стимулирует образование активной формы фосфорилазы - фосфорилазы а и активацию фосфофруктокиназы-ключевого фермента гликолиза. Однако даже максимально усиленный анаэробный метаболизм не способен длительно защищать уже поврежденный гипоксический миокард. Очень скоро запасы гликогена истощаются, гликолиз замедляется вследствие внутриклеточного ацидоза, который ингибирует фосфофруктокиназу. [45]
Страницы: 1 2 3 4