Меченая глюкоза
Cтраница 1
Меченая глюкоза внедряется в гексозаны и уроновые кислоты, причем только в небольшой ее части происходят изменения порядка С в углеродном скелете. При образовании пентозанов глюкоза теряет 6 - й атом углерода, а остальной скелет ( от 1-го до 5-го углеродного атома) не меняется. Меченая ксилоза и галактоза являются менее эффективными предшественниками как для пентозанов, так и для гексозанов. [1]
Получение меченой глюкозы основано на быстром изотопном обмбне углерода карбонильной группы глюкозы на радиоактивный углерод формальдегида и осуществляется нагреванием водного раствора глюкозы с формальдегидом - Си. В присутствии инсулина скорость этой реакции значительно возрастает. [2]
Следовательно, сразу же после подкормки растения меченой глюкозой отношение C6 / Q должно быть меньше единицы, хотя при последующем уравновешивании оно вновь достигнет единицы. [3]
Конн и Свэн [75] изучали образование галловой кислоты из меченой глюкозы, ацетата или фенилаланина в листьях Geranium pyrenacium. Из всех трех предшественников лучшим является глюкоза. [4]
Равномерно меченные аминокислоты можно получить биосин-тетически с помощью превращения 14СО2 в равномерно меченую глюкозу ( через крахмал), используя ее в качестве питательной среды для дрожжевых клеток; гидролиз полученного белка приводит к смеси равномерно меченных составляющих аминокислот. [5]
Прибавление к гомогенату мышц фосфорилазо-киназы ведет к увеличению концентрации фосфорилазы-а в системе и вместе с тем полностью тормозит внедрение в гликоген меченой глюкозы. В отсутствие киназы и фосфорилазы-а внедрение глюкозы в гликоген заметно возрастает. [6]
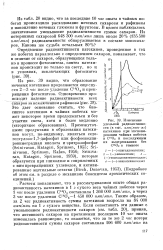 |
Изменение удельной радиоактивности ( в тыс. имп / мин-мг катехинов при экспонировании чайных побегов в С1402 и последующем их выдерживании без CuOj в темноте. [7] |
Из табл. 20 видно, что за последние 10 час опыта в чайных побегах происходило расходование меченых сахарозы и рафинозы и накопление меченых глюкозы и фруктозы. [8]
Исследованиями с меченой глюкозой древесины сосны Pinus strobus и Eucalyptus nitens / 27, 28 / был подтвержден механизм образования лигнина через шикимовую ( см. рис. 2) к префеновую ( рис. 3) кислоты. Аналогичные эксперименты с меченой шикимовой кислотой подтвердили, что этот механизм является при лигнификации решающим. Ацетатный механизм ( см. рис. 1) в процессе лигнификации не играет роли. Шикимовая кислота чрезвычайно распространена в высших растениях и в настоящее время установлено, что шикимовая и префеновая кислоты управляют синтезом ароматических соединений в микроорганизмах и растениях. На рис. 5 показано, что первичными ароматическими соединениями, образующимися по этому механизму, служат фенилпропаноиды - фенилпировиноградная ип-окси-фенилпировиноградная кислоты. Опыты с мечеными фенилаланином и тирозином однозначно доказали, что все растения обладают способностью инкорпорировать фенилаланин в ароматической части лигнина. С другой стороны, тирозин не играет такой универсальной роли в процессе лигнификации растений. Поэтому предпочтительным оказывается механизм с участием фенилапа-нина, превращаемого дезаминированием в коричную кислоту ( рис. 6), а не сходный механизм перехода тирозина в n - кумаровую кислоту. [9]
Одним из методов синтеза меченых соединений является биосинтез. Простейшим примером является получение меченой глюкозы и других углеводов в процессе фотосинтеза. Освещенные зеленые листья живого растения помещают в атмосферу меченной по углероду 14 ( С) углекислоты. Затем сахар экстрагируют из растений и очищают, используя обычные химические и биохимические процедуры. [10]
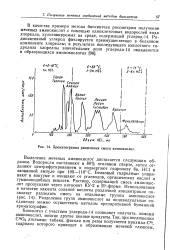 |
Хроматограмма разделения смеси аминокислот. [11] |
Биосинтез с участием углерода-14 позволяет получать, помимо аминокислот, многие другие ценные продукты. Так, при поглощении СиО2 листьями табака, фасоли или репы получается крахмал - С14, гидролиз которого приводит к образованию меченой глюкозы. [12]
В качестве ингибитора была использована глюкоза. В условиях, когда фермент, по всей вероятности, насыщен обычным для него акцептором, а именно водой, степень ингибирования ферментативной реакции глюкозой соответствует содержанию меченой глюкозы, вошедшей в глюкозо-б-фосфат. [13]
Так как каталитический участок активного центра р-амилазы расположен между вторым и третьим сайтами, на что указывают данные по составу продуктов ферментативного гидролиза ( почти исключительно мальтоза), то величины Л2 и Л3 не могут быть найдены с помощью описанного метода картирования активного центра. Поскольку мальтотриоза может связываться с активным центром р-амилазы лишь двумя продуктивными способами ( I и II, рис. 10), которые приводят соответственно к образованию меченых глюкозы и мальтозы как продуктов реакции, то можно заключить, что способ I - основной продуктивный способ связывания мальтотриозы. [14]
Создавалось впечатление, что в ткани мозга аминокислоты подавляют окисление глюкозы, а сами при этом окисляются. Выводы эти были основаны на следующих наблюдениях. В хвостовую вену крыс вводили 14С - глюкозу и через определенные промежутки времени прослеживали распределение изотопа в организме. В другой серии опытов срезы кор ы головного мозга инкубировали либо в присутствии одной лишь равномерно меченной глюкозы, либо в присутствии меченой глюкозы и немеченых глу-таминовой или аспарагиновой кислот. [15]
Страницы: 1 2