Is-элемент
Cтраница 1
IS-элементы содержат лишь те гены, которые необходимы для их собственного перемещения - транспозиции. [1]
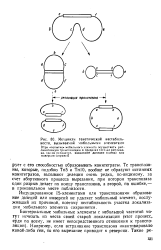 |
Механизм генетической нестабильности, вызываемой мобильными элементами. [2] |
Индуцированное IS-элементами или транспозонами образование делеции или инверсий не удаляет мобильный элемент, послуживший их причиной, поэтому нестабильность участка локализации мобильного элемента сохраняется. [3]
Именно поэтому из IS-элементов могли образоваться транспозоны. Любопытно проследить эволюцию составных транспозонов, в которых два IS-элемента окружают полезный ген. Довольно понятно, что эти структуры возникли в результате внедрения IS-элемента поблизости от этого гена и последующего перемещения того же элемента в другой участок молекулы ДНК по другую сторону от гена. Образовавшаяся структура может перемещаться как целое, но могут по отдельности перемещаться и образовавшие ее IS-элементы. Но если попавший между IS-элементами ген оказался удачным ( см. ниже), то получившийся транспозон начинает распространяться как целое, чему способствует постепенное накопление мутаций во фланкирующих его IS-элементах. Накопление мутаций приводит к дивергенции IS-элементов, причем по крайней мере один из них теряет способность к самостоятельной транспозиции. Так произошло, например, в случае транспозонов Тп5 и ТпЮ, лишь один IS-элемент которых способен к синтезу активной транспозазы и к самостоятельному перемещению. Транспозоны типа ТпЗ должны были возникнуть в результате более радикального процесса. [4]
Именно поэтому из IS-элементов могли образоваться транспозоны. Любопытно проследить эволюцию составных транспозонов, в которых два IS-элемента окружают полезный ген. Довольно понятно, что эти структуры возникли в результате внедрения IS-элемента поблизости от этого гена и последующего перемещения того же элемента в другой участок молекулы ДНК по другую сторону от гена. Образовавшаяся структура может перемещаться как целое, но могут по отдельности перемещаться и образовавшие ее IS-элементы. Но если попавший между IS-элементами ген оказался удачным ( см. ниже), то получившийся транспозон начинает распространяться как целое, чему способствует постепенное накопление мутаций во фланкирующих его IS-элементах. Накопление мутаций приводит к дивергенции IS-элементов, причем по крайней мере один из них теряет способность к самостоятельной транспозиции. Так произошло, например, в случае транспозонов Тп5 и ТпЮ, лишь один IS-элеменг которых способен к синтезу активной транспозазы и к самостоятельному перемещению. Транспозоны типа ТпЗ должны были возникнуть в результате более радикального процесса. [5]
Именно поэтому из IS-элементов могли образоваться транспозоны. Любопытно проследить эволюцию составных транспозонов, в которых два IS-элемента окружают полезный ген. Довольно понятно, что эти структуры - возникли в результате внедрения IS-элемента поблизости от этого гена и последующего перемещения того же элемента в другой участок молекулы ДНК по другую сторону от гена. Образовавшаяся структура может перемещаться как целое, но могут по отдельности перемещаться и образовавшие ее IS-элементы. Но если попавший между IS-элементами ген оказался удачным ( см. ниже), то получившийся транспозон начинает распространяться как целое, чему способствует постепенное накопление мутаций во фланкирующих его IS-элементах. Накопление мутаций приводит к дивергенции IS-элементов, причем по крайней мере один из них теряет способность к самостоятельной транспозиции. Так произошло, например, в случае транспозонов Тп5 и ТпЮ, лишь один IS-элемент которых способен к синтезу активной транспозазы и к самостоятельному перемещению. Транспозоны типа ТпЗ должны были возникнуть в результате более радикального процесса. [6]
 |
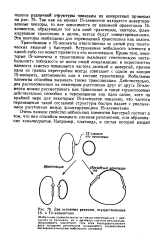
Две основные реакции, осуществляемые IS - и Тп-элементами. [7] |
Так как на концах IS-элементов находятся инвертированные повторы, то вне зависимости от взаимной ориентации IS-элементов, образующих тот или иной транспозон, повторы, фланкирующие транспозон в целом, всегда будут инвертированными. Эти повторы необходимы для перемещений транспозона как целого. [8]
 |
Две основные реакции, осуществляемые IS - и Тп-элементами. [9] |
Так как на концах IS-элементов находятся инвертированные повторы, то вне зависимости от взаимной ориентации IS-элементов, образующих тот или иной транспозон, повторы, фланкирующие транспозон в целом, всегда будут инвертированными. Эти повторы необходимы для перемещений транспозона как целого. [10]
 |
Две основные реакции, осуществляемые IS - и Тп-элементами. [11] |
Так как на концах IS-элементов находятся инвертированные повторы, то вне зависимости от взаимной ориентации IS-Элементов, образующих тот или иной транспозон, повторы, фланкирующие транспозон в целом, всегда будут инвертированными. Эти повторы необходимы для перемещений транспозона как целого. [12]
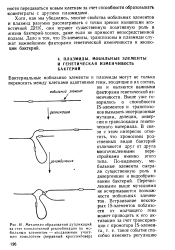 |
Механизм образования дупликаций за счет гомологичной рекомбинации по мобильным элементам - сподвижным участкам гомологии ( неравный кроссинговер. [13] |
Дело в том, что IS-элементы, транспозоны и плазмиды служат важнейшими факторами генетической изменчивости и эволюции бактерий. [14]
Выше уже говорилось о способности IS-элементов и транспозо-нов вызывать инсерционные мутации, делеции, инверсии и транслокации генетического материала. В связи с этим интересно отметить, что хромосомы родственных видов бактерий отличаются друг от друга многочисленными перестройками именно этого типа. [15]
Страницы: 1 2 3 4