Трансмембранный градиент
Cтраница 1
Трансмембранный градиент рН в хлоропластах создает электрохимический потенциал, обеспечивающий фосфорилирование. Ингибиторы фосфорилирования и так называемые разобщители ( среди них ионы аммония) могут уменьшить трансмембранный градиент рН из-за повышения проницаемости мембран хлоропластов, а не разрушать промежуточный X. [1]
Для поддержания столь высоких трансмембранных градиентов требуется энергия АТР. [2]
 |
Перенос ионов железа с помощью сидерофоров-гидроксаматов. [3] |
За счет этого трансмембранного градиента протонов и работают все виды вторичного транспорта. Протонный градиент в клетке всегда поддерживается на определенном уровне за счет дыхания, фотосинтеза, брожения. [4]
Эта схема показывает, как трансмембранный градиент концентрации ионов может быть сбалансирован градиентом напряжения. Внутри клетки концентрация К высокая, а снаружи низкая, тогда как концентрация Na снаружи выше, чем внутри. Здесь мембранные каналы свободно пропускают ионы К н не пропускают Na, поэтому распределение ионов К между двумя сторонами мембраны равновесное, а ионов Na - неравновесное. При равновесии электрохимический градиент К равен нулю; суммарный поток иоиов К тоже равен нулю. В результате устанавливается противоположно направленная ( по сравнению со случаем А) разность потенциалов, которая точно уравновешивает разность концентраций ионов Na по обе стороны мембраны. [5]
Использование световой энергии для создания трансмембранного градиента протонов происходит с участием бактериородо-псина и не связано с переносом электронов по цепи переносчиков. Этот хромопротеин с молекулярной массой 26 кДа содержит полипептидную цепь, построенную из 248 аминокислотных остатков и на 75 % состоящую из ос-спиральных участков. Ретиналь расположен параллельно плоскости мембраны и, следовательно, перпендикулярно белковым тяжам. [6]
Отвлечение части - Х на создание трансмембранного градиента рН могло закрепиться в процессе эволюции в связи с необходимостью обхода сравнительно медленной реакции - X - АТФ. Образование градиента Н не имеет лаг-фазы при смене темноты светом, в то время как образование АТФ. [7]
Градиенты концентрации между двумя сторонами клеточных мембран ( трансмембранные градиенты) варьируют очень сильно. Пожалуй, максимальный градиент концентрации в организме поддерживается плазматической мембраной обкладочных клеток слизистой оболочки желудка, секретирующих соляную кислоту в желудочный сок. По-видимому, эти клетки имеют какие-то очень активные мембранные насосы для секреции ионов водорода, так как для поддержания столь высокого градиента концентрации требуется значительное количество энергии. Перенос веществ через мембраны против градиента концентрации называют активным транспортом. Образование желудочной НС1 стимулируется особым, связанным с мембраной ферментом-т к называемой Н - транспортирующей АТРазой. [8]

Под влиянием индуцированного светом потока электронов возникает, следовательно, трансмембранный градиент концентрации ионов Н, так что с внутренней стороны тилакоидных пузырьков среда становится более кислой, чем с наружной. Все эти свойства согласуются с хемиосмотической гипотезой, предложенной первоначально для объяснения ркислительного фосфорилирова-ния, а позднее распространенной и на фотосинтетическое фосфорилирование. Схема на рис. 23 - 16 изображает поток ионов Н, движимых световой энергией; ионы Н 1 поступают из стромы внутрь тилакоида и вновь возвращаются наружу через молекулы АТР-синтетазы. В 1966 г. Андрэ Ягендорф поставил важный эксперимент, доказавший, что источником энергии для синтеза АТР действительно может служить трансмембранный градиент рН, обращенный щелочным концом наружу. [10]
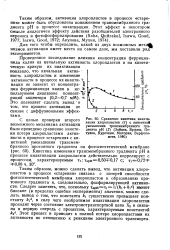 |
Сравнение кинетики инактивации хлоропластов ( 1 с кинетикой уменьшения трансмембранного градиента рН ( 2 ( Зайцев, Вржещ, Петухов, Курочкин, Маторин, Варфоломеев, 1980. [11] |
Таким образом, активация хлоропластов в процессе старения может быть обусловлена понижением трансмембранного градиента рН в процессе инактивации. [12]
В процессах дыхания и фотосинтеза освобождающаяся при переносе электронов энергия запасается первоначально в форме электрохимического трансмембранного градиента ионов водорода ( ДДН), т.е. имеет место превращение химической и электромагнитной энергии в электрохимическую. Последняя затем может быть использована для синтеза АТФ. Поскольку в обоих процессах синтез АТФ обязательно связан с мембранами, реакции, приводящие к его образованию, получили название мембранзави-симого фосфорилирования. Последнее подразделяется на два вида: окислительное ( АТФ образуется в процессе электронного переноса при окислении химических соединений) и ф о-тосинтетическое ( синтез АТФ связан с фотосинтетическим электронным транспортом) фосфорилирование. Таким образом, энергия, получаемая в процессах брожения, дыхания или фотосинтеза, запасается в определенных формах. [13]
Митчелла, энергия, освобождаемая в результате работы электронтранс-портной цепи, первоначально накапливается в форме трансмембранного градиента ионов водорода. [14]
Согласно этой гипотезе, градиент рН, который устанавливается меаду наружной и внутренней средой через мембрану хлоропласта ( трансмембранный градиент -), создает электрохимический потенциал, достаточный для синтеза АТФ. [15]
Страницы: 1 2 3