Трансмембранный градиент
Cтраница 2
Следует, однако, учесть, что до сих пор остается неясным вопрос о том, является ли трансмембранный градиент рН облигат-ным интера. АТФ в хлоропластах или таковым может быть и шсокоэнергетическ кое нефосфорилированное соединение Хе. [16]
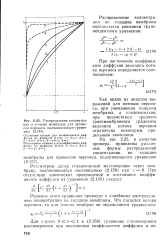 |
Распределения концентрации в сечении мембраны для процессов переноса, подчиняющихся уравнению. [17] |
Как видно из анализа выражений для потоков переноса, при уменьшении толщины мембраны, а следовательно, при возрастании среднего трансмембранного градиента величина потока переноса ограничена некоторым предельным значением. [18]
У морских ежей кратковременное повышение концентрации Са2 активирует специфические транспортные белки в плазматической мембране яйца ( возможно, при участии кальмодулина), которые используют энергию, запасенную в виде трансмембранного градиента иоиов Na, для откачивания ионов Н из клетки ( см. разд. Есть данные в пользу того, что именно это повышение рН индуцирует в оплодотворенных яйцах морского ежа позд. Во-вторых, если сразу после оплодотворения поместить яйца в морскую воду, не содержащую ионов Na ( так что не будет градиента Na4 для откачивания ионов Н), внутриклеточный уровень рН не повышается н поздние события, связанные с активацией яйца, не наступают. Такие яйца еще можно спасти, добавив к среде аммиака: тогда рН в клетке возрастает и даже при отсутствии внеклеточного Na индуцируется синтез белков и ДНК. [19]
Процесс образования карбоксисом для бактерий II фазы более редок, имеется развитая система мембран. АТФ также может образовываться за счет трансмембранного градиента протонов. Подсчитано, что для получения 1 моля НАДН необходимо окислить 5 молей нитрита с обратным переносом электронов. [20]
Когда этот градиент начинал уменьшаться вследствие выхода ионов Н из тилакоидов в среду, происходило образование АТР из ADP и фосфата. Поскольку АТР синтезировался в темноте, этот эксперимент доказывал, что трансмембранный градиент рН есть высокоэнергетическое состояние, посредством которого энергия переноса электронов передается АТР-синтетазе, получающей таким образом возможность генерировать АТР, как это постулируется хемиосмотической гипотезой. [21]
Трансмембранный градиент рН в хлоропластах создает электрохимический потенциал, обеспечивающий фосфорилирование. Ингибиторы фосфорилирования и так называемые разобщители ( среди них ионы аммония) могут уменьшить трансмембранный градиент рН из-за повышения проницаемости мембран хлоропластов, а не разрушать промежуточный X. [22]
Все живые организмы, включая простейших прокариот, отделены от окружающей среды клеточной мембраной. При этом роль мембран не ограничивается просто механическим отделением содержимого клетки. Они необходимы также для протекания некоторых основополагающих для живой материи процессов, в первую очередь производства АТФ. Как при окислительном фосфорилировании, так и при фотосинтезе сложный надмолекулярный комплекс, вмонтированный в мембрану, обеспечивает создание трансмембранного градиента концентрации протонов, за счет которого и происходит фосфорилирование АДФ. [23]
Под влиянием индуцированного светом потока электронов возникает, следовательно, трансмембранный градиент концентрации ионов Н, так что с внутренней стороны тилакоидных пузырьков среда становится более кислой, чем с наружной. Все эти свойства согласуются с хемиосмотической гипотезой, предложенной первоначально для объяснения ркислительного фосфорилирова-ния, а позднее распространенной и на фотосинтетическое фосфорилирование. Схема на рис. 23 - 16 изображает поток ионов Н, движимых световой энергией; ионы Н 1 поступают из стромы внутрь тилакоида и вновь возвращаются наружу через молекулы АТР-синтетазы. В 1966 г. Андрэ Ягендорф поставил важный эксперимент, доказавший, что источником энергии для синтеза АТР действительно может служить трансмембранный градиент рН, обращенный щелочным концом наружу. [24]
 |
Схема работы бакте. [25] |
Несмотря на кажущуюся простоту, очевидно, что бактериородопси-новая протонная помпа представляет собой сложную систему. Это означает, что поглощение кванта света должно приводить к возникновению напряженной конформации всего бактериородопсинового комплекса, служащей в дальнейшем источником энергии для переноса Н против электрохимического градиента. В организации такого переноса принимают участие ориентированные поперек мембраны а-спи-ральные тяжи и мембранные липиды, формирующие протонные каналы, природа и механизм действия которых пока не известны. Экстремально галофильные архебактерии содержат еще один ретинальбелковый комплекс - галородопсин, закачивающий на свету в клетки ионы хлора. Создающийся трансмембранный градиент СГ используется для синтеза АТФ. Одновременно этим обеспечивается также поддержание в цитоплазме высокой концентрации анионов, необходимой для уравновешивания высокой ионной силы внешней среды. [26]
Эта очень плодотворная идея дала сильный толчок развитию биоэнергетики за последние 35 лет. В своей хемоосмотической гипотезе Митчелл связывает протондвижущую силу с трансмембранной разницей электрохимических потенциалов, возникающих в водных фазах, разделенных мембраной. Включение в рассмотрение обширных водных фаз с необходимостью требует диссипации энергии, освобождаемой в ходе энергодонорного процесса ( электронный транспорт) по всему окружающему пространству. Отвергая эту точку зрения, Вильяме утверждает: Протоны в мембранах, а не осмотический трансмембранный градиент протонов требуются для обеспечения синтеза АТР. Он говорит также об энергиях локальных зарядов, генерируемых при отделении протонов от электронов соответствующими реакциями. [27]

Однако в соленых прудах, где концентрация NaCl иногда превышает 4 М, растворимость кислорода очень низка. Поэтому галофильным бактериям приходится иногда использовать другой источник энергии-солнечный свет. Они состоят из плотно упакованных молекул белка бактериородопсина ( мол. При освещении клеток молекулы бактериородопсина переходят в возбужденное состояние и временно обесцвечиваются. Когда затем эти возбужденные молекулы бактериородопсина, находящиеся в мембране, возвращаются в исходное основное состояние, выделяющаяся энергия используется на перекачивание ионов Н из клетки в наружную среду, вследствие чего возникает трансмембранный градиент рН, обращенный кислым концом наружу. Поскольку при этом концентрация ионов Н снаружи выше, чем внутри, эти ионы стремятся диффундировать обратно в клетку через находящиеся в мембране молекулы АТР-син-тезирующего фермента, напоминающего АТР-синтетазу митохондрий и хлоро-пластов. [29]
Перспективы развития мембранной технологии в большой мере связаны с надеждам. Уже сейчас промышленность располагает значительным набором мембран с селективными свойствами. Однако разработка и использование селективных мембранных материалов сталкивается до сих пор со значительными трудностями. Это связано главным образом с тем, что механизмы проницаемости как биологических, так и многих искусственных мембран окончательно не выяснены и не существует общего подхода к их описанию. Создание универсальной математической модели, адекватно описывающей мембранный транспорт, осложняется разнообразием процессов переноса через мембраны. В формировании реального процесса переноса могут принимать участие все механизмы в различных соотношениях. В силу специфических свойств мембран, больших трансмембранных градиентов и активного взаимодействия потока переноса со структурой мембраны наблюдаются значительные отклонения от закона Фика. При этом линейная зависимость потока переноса от градиента концентрации оказывается справедливой только для малых трансмембранных градиентов. Наблюдается замедление роста потока переноса или даже насыщение при больших значениях трансмембранного градиента. [30]
Страницы: 1 2 3