Тургорное давление
Cтраница 3
Как работают такие регуляторные системы. Эксперименты, проведенные с целью изучить быстрые реакции растительных клеток на изменения тургорного давления, показали, что тензодатчики, возможно, находятся в плазматической мембране. Так, например, внезапное понижение тургорного давления индуцирует активный перенос определенных молекул или ионов, чаще всего ионов К, внутрь клетки, тогда как повышение тургорного давления вызывает обратный эффект. Эти процессы протекают очень быстро и, по-видимому, связаны с какими-то изменениями в специфических транспортных белках плазматической мембраны. В отличие от этого образование осмотически активных молекул из запасных полимеров происходит более медленно. [31]

В вакуолях могут накапливаться и храниться самые различные молекулы, в том числе те необходимые для клетки вещества, которые потенциально опасны, если поступают в цитоплазму в больших количествах. Например, у некоторых растений в вакуолях специализированных клеток накапливаются такие хорошо известные продукты, как каучук и опиум. Даже вездесущие ионы Na могут селективно накапливаться в этих органеллах, где их осмотическая активность способствует поддержанию тургорного давления. Исследования, проведенные на клетках Nitella, показали, что натриевые насосы в то-нопласте поддерживают относительно низкую концентрацию Na в цитозо-ле за счет создания 4 - 5-кратного избытка зтих ионов в вакуолях. А поскольку вакуоль занимает в клетке Nitella гораздо больший объем, чем цитоплазма, получается, что основной пул клеточного натрия сосредоточен в вакуолях. [33]
Иногда создается неверное представление, что вода движется в листе по градиенту осмотического потенциала. Однако, хотя градиент водного потенциала существует, данных, которые позволили бы предположить, что осмотический потенциал в соответствующих клетках сильно различается, нет. Транспирационный ток поддерживается прежде всего за счет разности гидростатических потенциалов: потеря клеткой даже небольшого количества воды гораздо сильнее влияет на тургорное давление, чем на концентрацию растворенных веществ. То же самое можно сказать о корне ( разд. [34]
Клетки тканей животных не имеют обычно клеточной стенки. У клеток растений и многих микроорганизмов, напротив, имеется развитая многослойная клеточная стенка, находящаяся с наружной стороны от клеточной мембраны. Внутренние слои такой клеточной стенки служат конструкционным материалом, обеспечивающим достаточную жесткость формы клетки и устойчивость ее как к внешним механическим воздействиям, так и к тургорному давлению изнутри. [35]
Как работают такие регуляторные системы. Эксперименты, проведенные с целью изучить быстрые реакции растительных клеток на изменения тургорного давления, показали, что тензодатчики, возможно, находятся в плазматической мембране. Так, например, внезапное понижение тургорного давления индуцирует активный перенос определенных молекул или ионов, чаще всего ионов К, внутрь клетки, тогда как повышение тургорного давления вызывает обратный эффект. Эти процессы протекают очень быстро и, по-видимому, связаны с какими-то изменениями в специфических транспортных белках плазматической мембраны. В отличие от этого образование осмотически активных молекул из запасных полимеров происходит более медленно. [36]

Вместе с тем основные принципы построения всех клеточных стенок поразительно сходны, прочные волокна целлюлозы погружены в сложный трехмерный полисахаридный матрикс, который образуют связанные между собой молекулы пектинов и гемицеллюлозы. Эта так называемая первичная клеточная стенка способна выдерживать большие механические нагрузки и проницаема лишь для относительно небольших молекул. В водной среде растительная клетка, лишенная клеточной стенки ( протопласт), интенсивно поглощает воду путем осмоса, набухает и лопается. Величина тургорного давления весьма тонко регулируется, так как этот параметр очень важен для роста клеток и для обеспечения механической жесткости тканей молодого растения. [38]
Если клетка погружена в гипотонический раствор, то она поглощает воду со скоростью, пропорциональной разнице потенциалов воды внутри и вне клетки. Когда вода входит в клетку, содержимое клетки начинает оказывать на клеточную оболочку давление. Это гидростатическое давление называют тургорным давлением или тургором. [39]
После того как повилика присосется к питающему растению, ее связь с почвой нарушается и она начинает жить за счет питательных веществ, извлекаемых из растения-хозяина. При этом развивается паразит удивительно быстро, выбрасывая огромной длины желтоватые или оранжеватого цвета плети, в стеблевых узлах которых закладываются боковые побеги. Скоро растения в очаге заражения оказываются сильно опутанными длинными стеблями повилики. Вегетативное тело повилик обладает высоким тургорным давлением, которое позволяет обрывкам побегов не увядать в течение нескольких дней. [40]
В клетках растений резервные полисахариды могут находиться и в растворенном состоянии - в клеточном соке вакуолей. В этом соке обычно присутствуют фруктаны, а также сахароза и аналогичные олиго-сахариды, выполняющие, по-видимому, близкие биологические функции. Полисахариды, содержащиеся в вакуолярном соке, важны не только как резервы пищи организма. Их присутствие необходимо для поддержания так называемого тургорного давления - давления изнутри содержимого-клетки на клеточную стенку, что имеет первостепенное значение для всей жизнедеятельности растений. [41]
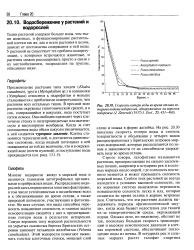 |
Скорость потери воды во время отлива четырьмя видами водорослей, обнаруженных на морском побережье ( J. Zaneveld ( 1937 J. Ecol, 25, 431 - 468. [42] |
Nymphaed) относятся к гидрофитам и меньше сталкиваются с проблемами осморегуляции, чем растения всех остальных типов. В пресной воде растения окружены гипотонической средой, и в сок, содержащийся в вакуолях, вода поступает путем осмоса. Она свободно проходит через клеточную стенку и полупроницаемые плазматическую и вакуолярную мембраны. Когда за счет поглощения воды объем вакуоли увеличивается, в ней создается тургорное давление. Клетка становится тургесцентной, и достигается равновесное состояние, когда водный потенциал в ней повышается до той же величины, что и в окружающей воде ( почти нулевой), и поступление воды прекращается ( см. разд. [43]
Несмотря на продолжающееся их движение через мембрану, изменения системы в целом прекратятся. Такое равновесие со средой называется динамическим. Осмотический потенциал клеточного содержимого останется скорее всего ниже, чем снаружи, поскольку для роста тургорного давления нужно не так много воды - меньше, чем для существенного разбавления внутреннего раствора. Однако эта разница компенсируется более высоким, чем снаружи, гидростатическим потенциалом клетки. Суммарные же водные потенциалы с обеих сторон клеточной стенки сравняются. [44]
В автолитический комплекс включают все деполимеразы, гидро-лизующие клеточные структуры и биополимеры. Первоначально к собственно автолизинам были отнесены те ферменты автолитиче-ского комплекса, которые участвуют в деградации материала клеточной стенки. У эубактерий - это ферменты, разрушающие пептидогли-кан ( ПГ) - гетерополисахарид, состоящий из остатков N-ацетилглюко-замина и N-ацетилмурамовой кислоты, образующих цепочки, соединенные пептидными мостиками. Фактически ПГ бактериальной стенки - это мешковидная макромолекула огромного размера, изолирующая протопласт, уравновешивающая его тургорное давление, придающая клетке форму и защищающая ее от внешних воздействий. [45]
Страницы: 1 2 3 4