Дефосфорилирование
Cтраница 2
Нуклеозиды обычно получают путем дефосфорилирования нуклеотидов при помощи препаратов соответствующих моно-эстераз ( стр. [16]
 |
Изменение обменной емкости и состава водных вытяжек при облучении в воде катионитов КФ-1 и КФ-Р. [17] |
Можно предположить, что кроме дефосфорилирования в полимерной матрице протекают процессы, аналогичные процессам, описанным для сульфокатионитов, которые приводят к образованию слабокислотных функциональных групп. [18]
АМФ оказывает влияние и на дефосфорилирование белков, к-рое осуществляется под действием протеинфосфатаз. Так, Протеинкиназа фосфорилирует один из белков, в результате чего он приобретает св-ва ингибировать фосфата-зу 1 - фермент, катализирующий дефосфорилирование. Метаболизм цАМФ осуществляют два фермента - аденил-атциклаза, катализирующая синтез цАМФ из АТФ, и фосфодиэстераза цАМФ, к-рая катализирует гидролиз цАМФ по З - фосфоэфирной связи с образованием 5 -адено-зинмонофосфата. [19]
Возможность работы мышцы за счет дефосфорилирования фосфагена была установлена в 1930 г. Лундсгардом. Этот автор установил, что мышца, отравленная монойодуксусной кислотой и таким путем лишенная способности расщеплять углеводы ( стр. После исчерпания запасов фосфагена и АТФ отравленная монойодацетатом мышца быстро утрачивает в анаэробных условиях способность отвечать сокращением на электрическое раздражение. [20]
Возможность работы мышцы за счет дефосфорилирования фосфагена была установлена в 1930 г. Лундсгардом. Этот автор установил, что мышца, отравленная монойодуксусной кислотой и таким путем лишенная способности расщеплять углеводы ( стр. При этом наблюдается распад фосфокреатина в большем размере, чем в неотравленной мышце. После исчерпания запасов фосфагена и АТФ отравленная монойодацетатом мышца быстро утрачивает в анаэробных условиях способность отвечать сокращением на раздражение с нерва. [21]
Последний подвергается при участии фруктозобисфофатазы дефосфорилированию в положении 1 с образованием седогептулозо-7 - фосфата. Эта реакция гидролиза фосфорного эфира необратима и дает возможность регулировать метаболизм в данном пункте. [22]
Реакция образования комплекса сопряжена с дефосфорилированием АТФ до АДФ. [23]
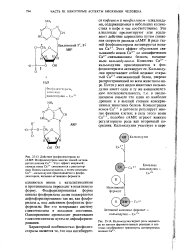
Фосфорилированная форма киназы фосфорилазы далее подвергается дефосфорилированию так же, как фосфо-рилаза а, под действием фосфатазы фосфорилазы. Все это возвращает систему гликогенолиза в исходное состояние. Одновременно происходит реактивация гликоген-синтазы путем ее дефосфорили-рования. [25]
Этот фермент может быть также использован для дефосфорилирования фос-фатидилэтаноламина и фосфатидилсерина [630], хотя эта реакция проходит с меньшей скоростью, чем в случае фосфатидил-холина. Нейтральные липиды, получаемые из фосфолипи-дов, анализируют, как описано выше в разд. [26]
Для выяснения причин пострадиационного падения вязкости и дефосфорилирования ДНК были проверены предположения о роли перекиси водорода, получающейся при радиолизе воды и лабильных продуктов, образующихся в цепи самой ДНК. Оказалось, что падение вязкости облученной ДНК наблюдается в условиях, исключающих действие перекиси водорода. [27]
Этот фермент катализирует, в частности, дефосфорилирование транскрибцион-ного фактора NF-AT, что в конечном итоге приводит к активации генов, с которых происходит считывание. [28]
Активность ГМГ-КоА-редуктазы регулируется также гормонально путем фосфорилирования - дефосфорилирования. Фосфорилированная форма ре-дуктазы неактивна, а следовательно, инсулин ее активирует, а глюкагон переводит в неактивную форму. Глюкокортикоиды ингибируют синтез редуктазы на уровне транскрипции гена этого фермента. Следует отметить, что синтез редуктазы в течение суток значительно варьирует, его максимум приходится на полночь, а минимум - на утренние часы. [29]
Ренконен [180], анализируя глицерофосфатиды, сначала проводил дефосфорилирование обработкой смесью уксусной кислоты и уксусного ангидрида. Полученные таким образом ди-глицеридацетаты он фракционировал по числу двойных связей в молекуле на слоях кремневой кислоты, пропитанных раствором нитрата серебра. [30]
Страницы: 1 2 3 4