Исследуемый - белка
Cтраница 2
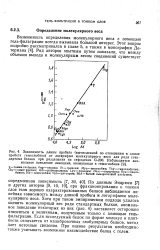
По данным Эндрюса [7] и других авторов [9, 10, 19], при фракционировании в тонком слое геля хорошо охарактеризованных белков наблюдается линейная зависимость между длиной пробега и логарифмом молекулярного веса. Хотя при таком эмпирическом подходе удалось оценить молекулярный вес ряда стандартных белков, аномальное поведение некоторых из них ( рис. 4) заставляет осторожно относиться к величинам, полученным только с помощью гель-фильтрации. Если исследуемый белок по форме молекул и плотности отличается от калибровочных глобулярных белков, метод дает неточные сведения. При оценке результатов необходимо также учитывать возможное взаимодействие белка с гелем. [17]
Экспериментально постоянную диффузии определяют путем измерения скорости размывания четкой горизонтальной границы, созданной в вертикальной трубке между белковым раствором и растворителем. Если исследуемый белок гомогенен, ход изменения градиента концентрации dc / dx будет соответствовать кривой распределения Гаусса, в то время как концентрационный градиент белковой смеси будет отклоняться от этой кривой. Скорость диффузии измеряют тогда, определяя то количество белка, которое прошло через диск в воду ( фиг. При помещении более тяжелого белкового раствора над растворителем диффузия ускоряется силой тяжести. Диффузионный сосудик калибрируется при помощи раствора, коэффициент диффузии которого известен. [18]
Если один и тот же белок обнаруживает несколько типов активности, то отношение этих активностей не должно изменяться в процессе очистки. Если же оно изменяется, то следует сделать вывод, что эти активности принадлежат разным белкам. Исследуемый белок не может считаться чистым, пока посторонние активности не будут устранены. [19]
После этого молекулярную массу и стоксов радиус исследуемого белка определяют путем интерполяции. Если исследуемый белок обладает какими-либо характерными свойствами, например ферментативной активностью или поглощением при определенной длине волны, его содержание в анализируемом препарате может быть минимальным. [20]
Когда в 1948 г. Мур и Стейн [55] в дополнение к классическим методам органической химии, а также манометрическому и бактериологическому анализу ввели ионообменную хроматографию, наступил поворотный момент в развитии химии аминокислот. В основу работы созданных сотрудниками Рокфеллеровского института современных автоматических аминокислотных анализаторов была положена ионообменная хроматография. Принцип работы этих приборов заключается в следующем. Исследуемый белок гидролизуют, затем гидролизат подвергают хроматографии на смоле типа дауэкс 50 х8 в Na-форме. Элюирование производят с помощью непрерывной подачи буферного раствора. Выходящий из колонки элюат попадает в пластмассовую ячейку особой формы, где он смешивается с раствором нингидрина. Подачу нингидрина осуществляет специальный насос, работающий синхронно с насосом, подающим буферный раствор на колонку. Затем смесь элюата с нингидрином проходит через тефлоновый капилляр, который погружен в кипящую баню. В этих условиях в растворах происходит нингидриновое окрашивание, интенсивность которого измеряется в проточной кювете спектрофотометрически. Поглощение света регистрируется самописцем. С помощью этой методики в одной и той же пробе можно определить кислые, нейтральные и основные аминокислоты, что не только экономит исследуемый материал, но и повышает точность и сокращает время исследования. [21]
Из изложенного видно, что определение плотности белковых растворов не представляет особых затруднений. Трудности возникают, однако, при измерении плотности сухих белков или влажных белковых препаратов. Многие белки при высушивании денатурируются, что делает невозможным использование этих препаратов для исследования гидратации. Это затруднение можно было бы преодолеть, определив предварительно плотность влажных кристаллов или раствора белка, а затем уже высушив белок. Однако при этом остается основная трудность, связанная с определением плотности сухого или влажного белка. Один из методов, применяемых для этой цели, состоит в том, что исследуемый белок помещают в пикнометр, взвешивают его, а затем наполняют пикнометр жидкостью известной плотности. Разумеется, такой заполняющей жидкостью не может служить вода, так как вода будет увеличивать степень гидратации; нельзя применять также этанол, ацетон и эфир, так как они способны притягивать воду и, следовательно, будут уменьшать степень гидратации. Обычно используют для этой цели бензол, бромбензол и другие гидрофобные жидкости. Но даже и при этом условии полученные результаты не вполне удовлетворительны, так как, во-первых, на поверхности раздела вода-растворитель имеет место денатурация белка, а во-вторых, небольшое количество воды все же растворяется в этих растворителях. Кроме того, при суспендиро-вании сухого белка в органической жидкости появляются маленькие пузырьки воздуха, снижающие точность определения. [22]
В качестве метода разделения для той же цели широко используют электрофорез в гелях. Для белков вследствие различий в их аминокислотном составе простой корреляции между молекулярной массой и подвижностью в электрофорезе не существует. Однако она может быть получена, если вести электрофорез в присутствии детергента. При разделении в SDS-полиакриламидном геле происходит полная денатурация белковых молекул с выворачиванием наружу их гидрофобного ядра в результате захвата додецильных радикалов, в результате чего на поверхности образовавшегося агрегата появляется отрицательный заряд и он ведет себя в электрофорезе как полианион. Эксперимент показывает, что при этом в широком диапазоне выполняется линейная зависимость электрофоретической подвижности от логарифма молекулярной массы, которая после градуировки по белкам с известной молекулярной массой может быть использована для определения молекулярной массы исследуемого биополимера. В результате денатурации субъединичные белки диссоциируют на отдельные субъединицы, так что, во-первых, выявляется, из скольких разнотипных субъединиц состоит исследуемый белок, и, во-вторых, находится масса отдельных субъединиц. [23]
Одну-две капли раствора аминокислот наносят на левый нижний угол такого листа и затем прилегающий к углу край погружают в растворитель, например в бутиловый спирт. Опустите край промокательной бумаги в воду, и вы увидите, как это происходит. Растворитель дойдет до того места, куда были нанесены аминокислоты, и потянет их за собой выше по бумаге. Каждое такое пятно может содержать не одну аминокислоту, а две или три. Для того чтобы их также разделить, бумагу высушивают, поворачивают на 90 градусов и уже другую, смежную, сторону листа погружают в другой растворитель, который разделит компоненты, присутствующие в пятнах аминокислот. В конце процедуры бумагу высушивают и сбрызгивают реактивом, который окрашивает аминокислоты, теперь они видны на бумаге как окрашенные пятна. Зрелище получается довольно эффектным: все аминокислоты, которые находились в одном растворе, разошлись по бумаге в вертикальном и горизонтальном направлениях и образовали мозаику из цветных пятен. Опытные биохимики по положению пятна могут сказать, какая аминокислота в нем находится, и, следовательно, им достаточно одного взгляда, чтобы сказать, из каких аминокислот состоит исследуемый белок. Вырезав пятно и вымыв из бумаги аминокислоту, они могут определить, сколько ее содержалось в данном белке. [24]
Страницы: 1 2