Ферментативный аппарат
Cтраница 3
 |
Схема репликации генома SV40 ( схема Кэрнса. [31] |
После взаимодействия вирус-специфической хеликазы - большого Т - антигена - с участком ori в кольцевой двухнитевои ковалентно-непрерывнои молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - ( гации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 6-типа или 6-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент-йую непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [32]
 |
Схема репликации генома SV40 ( схема Кэрнса Жирные лиинн - родительские цепи ДНК. тонкие - дочерние. [33] |
После взаимодействия вирус-специфической хеликазы - большого Т - антигена - с участком on в кольцевой двухнитевой ковалентно-непрерывной молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - [ ации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 6-типа или 9-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент - Ную непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [34]
 |
Схема репликации генома SV40 ( схема Кэрнса Жирные линии - родительские цепи ДНК. тонкие - дочерние. [35] |
После взаимодействия ярус-специфической хеликазы - большого Т - антигена - с участком ori в кольцевой двухнитевой ковалентно-непрерывной молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - ации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 9-типа или 0-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент-йую непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [36]
Существование в хлоропластах ферментативного аппарата для фиксации двуокиси углерода было показано лишь 30 лет спустя Арноном и др. [1 ] в опытах с использованием С14 и значительно большего набора биохимических кофакторов, чем тот, который имелся в распоряжении Хилла. Эти опыты послужили основой для общепринятой в настоящее время концепции, согласно которой как световые, так и темновые реакции фотосинтеза протекают в хлоропластах. [37]
Почвенная микрофлора имеет хорошо развитую пищевую сеть и мощный компенсационный механизм, основанный на функциональной взаимозаменяемости одних видов другими. Кроме того, благодаря лабильному ферментативному аппарату многие виды могут легко переключаться с одного питательного субстрата на другой, обеспечивая тем самым стабильность экосистемы. Это существенно усложняет оценку воздействия на нее различных антропогенных факторов и требует использование интегральных показателей. [38]
Складывается впечатление о большой связи с показателями селекционного значения эмбриофизиологических и обменных процессов. На различных стадиях эмбриогенеза и в периоды усиленного роста поддерживающий их ферментативный аппарат обобщается и становится важным компонентом функциональной системы, обеспечивающей продуктивность. [39]
По составу ферментов облигатные метилотрофы разделяются на две группы. Те из них, которые имеют мембранную систему типа II, по ферментативному аппарату похожи на факультативные метилотрофы. Те же, которые имеют мембранную систему типа I, обладают набором ферментов, который аналогичен набору ферментов многих облигатных хемоавтотрофов. [40]
Вначале репликация вдет по схеме Кэрнса: обнаруживаются 6-молекулы, в которых две репликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [41]
Вначале репликация идет по схеме Кэрнса: обнаруживаются 9-молекулы, в которых две репликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [42]
Вначале репликация идет по схеме Кэрнса: обнаруживаются 6-молекулы, в которых две уепликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [43]
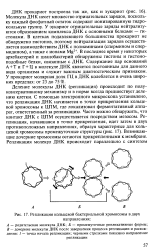 |
Репликация кольцевой бактериальной хромосомы в двух. [44] |
Деление молекулы ДНК ( репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Возникающие дочерние хромосомы остаются прикрепленными к мембране. [45]
Страницы: 1 2 3 4