Карбамоилфосфат
Cтраница 2
 |
Орнитиновый цикл синтеза мочевины в печени. [16] |
Из приведенной схемы процесса мочевинообразования нетрудно видеть, что один из атомов азота мочевины имеет своим источником свободный аммиак ( через карбамоилфосфат); второй атом азота поступает из ас-партата. Аммиак образуется главным образом в процессе глутаматде-гидрогеназной реакции. В процессе пополнения запасов аспартата участвуют три сопряженные реакции: сначала фумарат под действием фумаразы присоединяет воду и превращается в малат, который окисляется при участии малатдегидрогеназы с образованием оксалоацетата; последний в реакции трансаминирования с глутаматом вновь образует аспартат. [17]
Для объяснения этих фактов исследователи группы Линена предположили, что данный микроорганизм образует биотин из цистеина, пимелил - КоА и карбамоилфосфата ( фиг. [18]
Можно лишь предположить, что резкое пострадиационное уменьшение внутриклеточной концентрации АТФ, обнаруживаемое в кроветворных тканях облученных животных, должно снизить интенсивность этой реакции и, как следствие, уменьшить содержание в клетке карбамоилфосфата, являющегося субстратом аспарат - карбамоилтрансферазы - второго фермента данного пути биосинтеза УМФ. Однако в относительно радиорезистентной ткани - печени - концентрация карбамоилфосфата в течение первых 4 сут после рентгеновского облучения мышей в дозе 178 мКл / кг практически не изменяется. Достоверное уменьшение фонда этого соединения отмечается только на 6 - 8 - е сутки лучевой болезни. Через 9 сут его уровень нормализуется, а через 10 сут начинает резко возрастать и к концу 2 - й недели лучевой болезни уже в 2 раза превышает контрольное значение [ Waldschmidt M. Следовательно, в печени мышей на протяжении первых 5 сут после лучевого поражения 1 - й этап биосинтеза УМФ de novo не нарушается, затем, по-видимому, несколько угнетается и значительно усиливается через 12 - 14 сут. [19]
Аммиак входит в состав органических соединений посредством основных реакций, в которых он связывается: а) в виде а-амино-группы, б) в виде амидных групп аспарагина и глутамина, в) в виде карбамоилфосфата - промежуточного продукта в синтезе цитрул-лина ( см. стр. [20]
Торможение глутаминсинтетазы карбамоилфосфатом, которое у позвоночных, видимо, составляет неотъемлемую часть механизма, регулирующего обмен NH, у моллюсков не могло бы быть выгодной особенностью ( если только у них нет других ре-гуляторных изоферментов глутаминсинтетазы), так как блокада в этом пункте препятствовала бы также дальнейшему синтезу карбамоилфосфата. [21]

Карбамоилфосфат-синтетаза I представляет собой регуляторный фермент; положительным, или активирующим, модулятором служит для нее N-ацетилглу-тамат. Карбамоилфосфат ( рис. 19 - 19) принадлежит к высокоэнергетическим соединениям; его можно считать активированным донором карбамоильных групп. Отметим, что при образовании одной молекулы карбамоилфосфата используются концевые фосфатные группы двух молекул АТР. [23]
Далее карбамоилфосфат взаимодействует с орнитином ( второй фермент - орнитинкарбамо-илтрансфераза) с образованием цитруллина: карбамоилфосфат ор-нитин - цитруллин. [24]
Биосинтез пиримидинов начинается с синтеза карбамоилфосфата, промежуточного продукта, важного и в других отношениях. Из всех известных путей образования карбамоилфосфата наибольшее значение имеет, безусловно, синтез его из аммиака, двуокиси углерода и АТФ. Известны два типа таких реакций. [25]
Общая схема синтеза мочевины из двуокиси углерода и аммиака представлена на фиг. Первый этап этой последовательности реакций - образование карбамоилфосфата - является в то же время и первым этапом синтеза пиримидинов ( см. гл. Орнитинтранскарбамилаза ( карба-моилфосфат: L-орнитин - карбамоилтрансфераза) катализирует конденсацию карбамоилфосфата и орнитина с образованием цитруллина. Этот фермент обнаруживается в печени млекопитающих, у высших и низших растений и у некоторых микроорганизмов. У птиц он отсутствует. Превращение цитруллина в аргинин осуществляется в результате двух последовательно протекающих реакций, чрезвычайно сходных с реакциями, приводящими к синтезу адениловой кислоты ( см, гл. [26]
Так, пять ферментов осуществляют биосинтез уридин-фосфата из карбамоилфосфата и аспартата. Последние четыре фермента этого процесса репрессируются координированно. [27]
Можно лишь предположить, что резкое пострадиационное уменьшение внутриклеточной концентрации АТФ, обнаруживаемое в кроветворных тканях облученных животных, должно снизить интенсивность этой реакции и, как следствие, уменьшить содержание в клетке карбамоилфосфата, являющегося субстратом аспарат - карбамоилтрансферазы - второго фермента данного пути биосинтеза УМФ. Однако в относительно радиорезистентной ткани - печени - концентрация карбамоилфосфата в течение первых 4 сут после рентгеновского облучения мышей в дозе 178 мКл / кг практически не изменяется. Достоверное уменьшение фонда этого соединения отмечается только на 6 - 8 - е сутки лучевой болезни. Через 9 сут его уровень нормализуется, а через 10 сут начинает резко возрастать и к концу 2 - й недели лучевой болезни уже в 2 раза превышает контрольное значение [ Waldschmidt M. Следовательно, в печени мышей на протяжении первых 5 сут после лучевого поражения 1 - й этап биосинтеза УМФ de novo не нарушается, затем, по-видимому, несколько угнетается и значительно усиливается через 12 - 14 сут. [28]
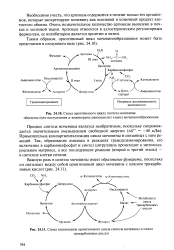 |
Схема орнитинового цикла синтеза мочевины. обведены пути поступления а-аминогрупп аминокислот в цикл мочевинообразования.| Схема взаимосвязи орнитинового цикла синтеза мочевины и цикла. [29] |
Примечательна компартментализация цикла мочевины и связанных с ним реакций. Так, образование аммиака в реакциях трансдезаминирования, его включение в карбамоилфосфат и синтез цитруллина происходят в митохонд-риальном матриксе, а все последующие реакции ( второй и третий этапы) - в цитозоле клетки печени. [30]
Страницы: 1 2 3 4