Электронно-микроскопическая картина
Cтраница 2
К сожалению, по этим данным трудно установить особенности связи между содержанием ДОФ и размерами структурных образований, однако видно, что существует отчетливая тенденция к увеличению размеров по мере возрастания содержания ДОФ. Джейл отмечает, что отсутствие строгой системы в полученных им электронно-микроскопических картинах можно объяснить влиянием условий получения образцов. [16]
С целью выяснения причин возникновения пористой структуры в частице лесса в водных суспензиях была исследована при помощи электронного микроскопа картина строения частиц лессовых суспензий в воде, затем вода в суспензии заменялась этиловым спиртом. Мы считали возможным проводить обезвоживание лесса этиловым спиртом вследствие того, что электронно-микроскопическая картина частиц лесса, полученных распылением и из спиртовых суспензий, совершенно идентична. [17]
Процессы старения и связанные с ними изменения формы и строения коллоидных частиц в золях сернистого мышьяка протекают очень медленно. Электронно-микроскопическая картина отличается лишь тем, что в старых золях можно наблюдать большее количество агрегатов. [18]
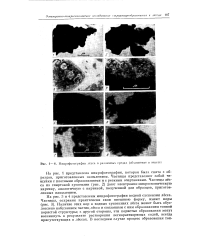 |
Микрофотографии лесса в различных средах ( объяснение в тексте. [19] |
На рис. 1 представлена микрофотография, которая была снята с образцов, приготовленных напылением. Частицы представляют собой чешуйки с плотными образованиями и с резкими очертаниями. Частицы лесса из спиртовой суспензии ( рис. 2) дают электронно-микроскопическую картину, аналогичную с картиной, полученной для образцов, приготовленных напылением. [20]
 |
Схематическое изобра-жение образовавшейся в сплошном материале микротрещины, пронизанной тяжами. [21] |
При растяжении тонких пленок деформация развивается резко неоднородно с формированием начальных дефектов размером порядка 200 А, которые, сливаясь, образуют субмикротрещины в направлении, перпендикулярном оси ориентации. Микротрещины, возникающие в низкомолекулярных образцах, вполне подобны обычным макроскопическим разрывам и представляют собой просто пустоты. Но субмикротрещины, образовавшиеся в высокомолекулярных образцах, выглядят совершенно по-иному, и электронно-микроскопическая картина субмикротрещины оказывается аналогичной схеме, показанной на рис. VI.7. Образование субмикротрещины, пересеченной фибриллизованным материалом, происходит при переходе к молекулярным весам порядка 4 - Ю4, что хорошо согласуется с характерными значениями молекулярных весов, при которых появляется сетка зацеплений макромолекул в полистироле. [22]
При студнеобразовании в жестких условиях, когда скорость установления равновесного состава фаз очень велика, внутренние напряжения, возникающие в матричной фазе, не успевают рассасываться, и происходит частичное нарушение сплошности этой фазы. В результате этого отдельные микроучастки низкоконцентрированной фазы сливаются. Происходит внутренний микросинере-зис, что приводит к укрупнению элементов структуры, увеличению отражающих поверхностей и изменению электронно-микроскопической картины студней. Под влиянием внутренних напряжений может происходить частичная деформация элементов матричной фазы, и они приобретают анизометрию, становясь некоторым подобием фибриллярных образований. [23]
На рис. 5 приведены электронно-микроскопические снимки, полученные с серебряных частиц после электродиализа желатинового слоя. Приведенные фотографии явно указывают на аморфную структуру частиц при небольшой продолжительности созревания эмульсий ( до 8 час. К сожалению, с полученных препаратов не удалось получить четких электроно-грамм, что объясняется небольшим числом крупных частиц в поле зрения, однако электронно-микроскопическая картина здесь настолько ясна, что сама по себе в достаточной степени характеризует процесс. [24]
Считая, что в реакции принимают участие указанные вещества, Келвин предложил схему энергетически приемлемого процесса. Восстановленная липоевая кислота сама восстанавливает три-фосфопиридинуклеотид в ТПН-Н, а последний вызывает реакцию, являющуюся критической ступенью процесса фотосинтеза-восстановление фосфоглицериновой кислоты в триозофосфат. Таким образом, основная функция хлорофилла в процессе фотосинтеза состоит в том, что он играет роль вещества, разделяющего заряды. Исходя из полученной Шестрандом электронно-микроскопической картины правильного слоистого строения гранул и хлоропластов, содержащих хлорофилл [7], Келвин пришел к заключению, что эта слоистая структура представляет собой как бы фотобатарею. Таким образом, электрон достигает слоя, содержащего тиол, в то время как на противоположном полюсе фотобатареи происходит разложение воды. Слоистое строение гранул хлоропластов очень напоминает структуру наружных члеников палочек и колбочек в органах зрения живых организмов ( см. гл. Было бы очень важно установить, можно ли при помощи микроэлектродной техники обнаружить фототоки в хлоропластах, так же как это уже было сделано для сетчатки. [25]
 |
Электронно-микроскопические фотографии ( получены по методу реплик кристаллов, образовавшихся при полимеризации этилена из газовой фазы в присутствии TiCl3 - AlClEt2 в качестве катализатора. [26] |
На основании изложенных соображений можно предположить, что кристаллы с выпрямленными цепями не могут образовываться просто в результате непрерывного отложения сегментов полимера. Наблюдения с помощью электронного микроскопа показали, что продукты полимеризации как в случае синтеза полиметилена из ди-азометана ( см. рис. III.96), так и при получении полиэтилена радиационной полимеризацией ( см. рис. III.97) представляют собой смесь большого числа ламелярных кристаллов и перепутанных волокон. Пример такой структуры показан на рис. III.98, полученном с помощью метода реплик. Как показали исследования образцов, подвергнутых обработке азотной кислотой, наблюдаемые на электронно-микроскопических картинах волокнистые образования представляют собой несовершенные кристаллы. Это наглядно видно из рис. II 1.99, на котором наблюдаются оставшиеся после обработки ламели и совершенно отсутствуют волокнистые структуры. [27]
На снимке вискозного студня ( пленки), не подвергшегося ориентации ( рис. 11.11, а), отчетливо видны отдельные анизометрические образования, составляющие своеобразную непрерывную сетку. На рис. 11.11, б показан результат одноосного растяжения такой застудневающей пленки. На нем отчетливо видна ориентация указанных фазовых образований вдоль направления действия растягивающего усилия. Полученная структура близка к конечной структуре готового волокна, что видно из рис. 11.11, в, на котором приведена электронно-микроскопическая картина скола вискозного волокна. Как видно из сравнения снимков, фазовые анизометрические образования, возникающие при проведении начальной стадии формования, и структурные элементы в готовом волокне имеют сходную форму и размеры. Такое совпадение, правда, еще не может служить окончательным доказательством того, что конечная структура волокна начинает зарождаться уже на первой стадии фазовых преобразований раствора полимера. Однако не исключено, что именно этим обстоятельством объясняется способность к макрофибриллизации при механической дезинтеграции сухих или набух-тих волокон, которые были сформованы путем осаждения из раствора полимера. [28]
Действительно, хорошо известно, что кристаллический полимер растворяется гораздо хуже, чем аморфный полимер того же химического строения. Ориентированные образцы также хуже растворяются по сравнению с изотропными образцами. Возможно, что и в случае изотропных образцов аморфных полимеров надмолекулярная структура может быть различной, однако этот вопрос до сих пор является предметом дискуссии. Трудно, однако, представить, что если поверхность пленки, полученной из раствора, и поверхность скола блочного образца, полученного из расплава, дают одну и ту же электронно-микроскопическую картину глобул, то эта картина является следствием артефактов. [29]
Более половины валового синтеза РНК эукариотической клетки приходится на образование 18S, 28S и 5 85 РНК рибосом. Рибосо-мальные гены представлены сотнями повторяющихся копий, сгруппированных в одном или нескольких участках генома. Многократная повторяемость генов рРНК, а также их амплификация на определенных стадиях развития отражают потребность клетки в больших количествах транскриптов этих генов. Ранее считали, что районы спейсера не транскрибируются. Действительно, электронно-микроскопические картины транскрипции рибосомных генов, казалось бы, подтверждали этот вывод. Однако оказалось, что транскрипты спейсеров подвергаются особенно быстрому процессингу и деградации. В конце спейсера находится сайт терминации Т, включающий консервативную последовательность из нескольких нуклеотидов. Функциональная особенность этого сайта заключается в том, что он же отвечает и за реинициацию нового транскрипта. Одно и то же мутационное изменение в этом районе спейсера снижает как эффективность терминации, так и реинициации транскрипции. При освобождении транскрипта в сайте Т полимераза сохраняется связанной с ДНК. Взаимодействуя с дополнительным белковым фактором, полимераза тут же настраивается на следующий акт реинициации транскрипции. Благодаря сопряжению актов терминации и инициации в одном сайте обеспечивается высокая эффективность процесса транскрипции, поскольку каждый новый акт инициации не требует поиска промотора. [30]
Страницы: 1 2 3