Киназа
Cтраница 2
Перенос конечного фосфатного остатка АТФ на различные акцепторы катализируется специфическими ферментами киназами. Можно считать, что механизм этой реакции заключается в нуклеофильной атаке на конечный фосфат АТФ. [16]
Приведенные выше характеристики ферментов биосинтеза ТМФ в полной мере относятся и к киназам, фосфорилирующим его до трифосфата. Так, например, отмечено, что экстракт нормальной печени крысы почти не фосфорилирует ТМФ, хотя остальные дезоксимононуклеотиды фосфорилируются нормально. Установлено, что киназы, катализирующие биосинтез ТТФ из тимидина или его монофосфата, отсутствуют в нормальной, но появляются в регенерирующей печени крысы и в других тканях, характеризующихся быстрой скоростью роста. К этому моменту в регенерирующей печени сильно увеличивается активность ДНК-полимеразы - фермента заключительного этапа синтеза ДНК. [17]
Механизм образования фермент-субстратного комплекса удобно проследить на реакциях, катализируемых фосфомутазами или киназами. Одним из этих ферментов является фосфоглкжо-мутаза, катализирующая равновесие между глкжозо-1 - фосфатом и глюкозо-6 - фосфатом. Фермент содержит прочно связанный фосфор, который нельзя удалить диализом. [18]
Марганец выполняет функцию активатора целого ряда ферментов, относимых в первую очередь к киназам, кабоксилазам и трансфера-зам. В случае реакций, активируемых марганцем, ион металла вступает во взаимодействие с субстратом, содержащим фосфатный остаток ( особенно АТФ), образуя хелат, или реагирует непосредственно с белком. Марганец химически близок к магнию. Исходя из этого, активация большинства ферментативных реакций этим элементом носит неспецифичеокий характер и он может быть заменен магнием [ Keen С. В то же время действие Мп2 и Mg2 может быть совершенно различным. Так, оба иона образуют комплексы с АТФ. [19]
Конваллотоксин) 207 p - L-Рамнопиранозид строфантидина 207 L-Рамнулозофосфат - альдолаза 378 L-Рамнулозофосфат - киназа 378 Распад углеводов 363 ел. [20]
В реакциях первого типа осуществляется перенос высокоэнергетической фосфатной группы от молекулы-донора на АДФ, катализируемый соответствующими киназами. Реакциями такого типа являются реакции субстратного фосфорилирования на пути анаэробного превращения Сахаров. У прокариот, имеющих ЦТК, реакция превращения сукцинил - КоА в янтарную кислоту сопровождается запасанием энергии в фосфатной связи ГТФ, который затем отдает фосфатную группу АДФ. [21]
Неактивная фосфорилаза ( Ь) превращается в активную фосфорилазу ( а) в результате ее фосфорилирования фосфорилаза-6 - киназой. Однако последняя также существует в фосфорилированной ( активной) и нефосфорилированной ( неактивной) формах. Протеинкиназа может фосфорилировать не только фосфо-рилазокиназу, но также многие другие белки, такие как казеин и даже гликогенсинтетазу. Фосфорилирование гликогенсинтетазы инактивирует фермент, и в итоге сАМР не только активирует деградацию гликогена, но также и выключает его синтез. В результате каскадного процесса, описанного выше, эффект малого количества сАМР усиливается, сАМР может осуществлять очень тонкий контроль за процессом деградации гликогена. [22]
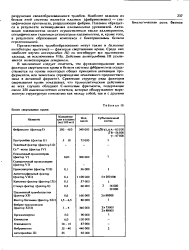 |
Белки свертывания кровн. [23] |
Активация плазминогена может осуществляться также калликреином, специфическим тканевым активатором плазминогена, и, кроме того, в результате образования комплекса с бактериальным белком стрепто киназой. [24]
Важная особенность этого переноса состоит в том, что почти все сверхвысокоэнерге-тические фосфорилированные соединения передают свои фосфатные группы низкоэнергетическим акцепторам фосфата через АТР, так что передача совершается в два этапа; оба эти этапа катализируются специфичными киназами. [25]
Сама по себе эта киназа, по-видимому, активна как в темноте, так и на свету; свет, вероятно, вызывает конфор-мационный переход родопсина из формы, которая не может быть фосфорилирована, в конформацию, узнаваемую киназой в качестве субстрата. В каждую молекулу обесцвеченного родопсина может входить 7 - 9 фосфатных групп. In vitro фосфорили-рование родопсина протекает медленно, реакция требует нескольких минут, если обесцвечено значительное количество-родопсина. При небольших уровнях обесцвечивания эта реакция протекает, однако, значительно быстрее. [26]
 |
Интегративная и амфиболическая функции цикла трикарбоновых кислот. [27] |
Как отмечалось выше, в состав ПД-комплекса входят регуляторные ферменты киназа и фос-фатаза. Киназа катализирует фосфорилирование ПДГ ( Ej) и переводит этот фермент в неактивное состояние, а фосфатаза отщепляет от него фосфорную кислоту и активирует его. [28]
В результате реакции АТР превращается a ADP. Киназа осуществляет также обмен 5 -концевых фосфатных групп олиго - или полинуклеотидов с у-фосфатной группой АТР в присутствии избытка ADP. Фермент используют для введения меченых фосфатных групп в 5 -концевые звенья ДНК. [29]
Лимитирующей стадией процесса трансляции является ее инициация. Реакция катализируется ферментом 1Р2 - киназой, причем присоединение фосфатной группы инактивирует фактор инициации. Этот феномен был изучен на примере синтеза гемоглобина в ретикулоцитах. Сначала было установлено, что глобин синтезируется только в присутствии гема. Затем была выстроена вся система регуляции синтеза глобина. Оказалось, что активация 1Р2 - киназы происходит за счет ее фосфорилирования цАМФ - зависимой протеинкиназой. Взаимодействие этой протеинкиназы с цАМФ и ее активацию блокирует гем, выполняя тем самым негативный контроль синтеза гемоглобина. [30]
Страницы: 1 2 3 4