Компартментализация
Cтраница 1
 |
Схема аллостерической регуляции активности фермента. [1] |
Компартментализация особенно распространена у эукариотических микроорганизмов, у которых много орга-нелл с собственными мембранами. Например, окисление жирных кислот локализовано в митохондриях, тогда как синтез жирных кислот происходит в цитоплазматическом матриксе. Компартментализация делает возможным одновременное, но раздельное регулирование анаболических путей. Далее активность данного пути может координироваться в общем метаболизме путем регуляции транспорта метаболитов и коферментов между клеточными ком-партментами. Неоднородность клеточной внутренней среды позволяет создавать градиенты метаболитов около определенных ферментных систем. [2]
Компартментализация прокариотической клетки значительно менее выражена, чем у эукариотических клеток. ДНК не окружена ядерной мембраной, а органеллы типа митохондрий и хлоропластов отсутствуют. Область ядра, видимая на электронной микрофотографии ультратонкого среза клетки в виде сетчатой структуры из тонких нитей, непосредственно граничит с заполненной рибосомами цитоплазмой. [3]
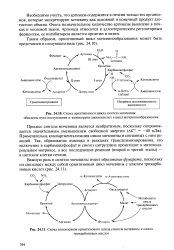 |
Схема орнитинового цикла синтеза мочевины. обведены пути поступления а-аминогрупп аминокислот в цикл мочевинообразования.| Схема взаимосвязи орнитинового цикла синтеза мочевины и цикла. [4] |
Примечательна компартментализация цикла мочевины и связанных с ним реакций. Так, образование аммиака в реакциях трансдезаминирования, его включение в карбамоилфосфат и синтез цитруллина происходят в митохонд-риальном матриксе, а все последующие реакции ( второй и третий этапы) - в цитозоле клетки печени. [5]
Теория компартментализации, согласно которой синтетические процессы в клетке пространственно отделены от процессов расщепления, находит себе подтверждение в метаболизме крахмала; мы имеем в виду фосфорилазную и амилазную реакции. Однако приведенные выше открытия по-новому раскрывают эти два различных механизма, имеющих место в клетке: 1) синтез протекает с участием АДФГ и 2) расщепление происходит при участии фосфорилазы. Природа, по-видимому, организовала в живой клетке независимую работу механизмов для синтеза и расщепления макромолекул; это относится и к крахмалу, и к белку, и к полинуклеотидам. [6]
Эволюционные преимущества компартментализации очевидны, они обсуждаются в последующих параграфах. [7]
Многотомное издание, содержащее обзорные статьи по компартментализации различных видов биохимической активности в клетках. [8]
Следовательно, эндогенное АДФ-рибозилирование могло бы регулировать степень компартментализации EF-2 на полирибосомах эукариотической клетки. Ассоциация Ef-2 с полирибосомами, вероятно, имеет несколько целей: во-первых, она поддерживает увеличенную локальную концентрацию EF-2 непосредственно вблизи мест его функционирования, что может быть важным в большом объеме эукариотической клетки; во вторых, избыточный EF-2, обратимо связанный с мРНК, возможно, имеет особую функцию в преодолении трансляционных барьеров, например, стабильной вторичной структуры. [9]
Другой интересный аспект результатов по перекрестной релаксации связан с компартментализацией воды в ткани. Так как скорости релаксации при перекрестной релаксации могут быть вычислены путем решения сопряженных дифференциальных уравнений, наблюдаемые скорости релаксации нельзя представить в виде простой суммы вкладов внутримолекулярной и перекрестной релаксации. В типичном случае на релаксацию растворителя оказывает существенное влияние перекрестная релаксация, даже если ее временная зависимость определяется одним мгновенным значением и поэтому может быть принята за экспоненциальную. Однако релаксация протонов белка ( если она наблюдается) почти всегда проявляется в форме двух экспоненциальных вкладов. Устранение слагаемого, описывающего перекрестную релаксацию, приводит к устранению и одного из этих вкладов, изменяя при этом величину другого, а также скорость релаксации протонов растворителя. Изменение релаксационного поведения, состоящее в переходе от процесса, описываемого с помощью двух экспонент, к процессу с единственной экспонентой, может быть ошибочно принято за замещение одного из двух типов протонов дейтронами. В тканях ситуация еще более сложна, но, как мы полагаем [15], большая часть результатов по протонной релаксации в образцах дейтерированных тканей нуждается в перепроверке в свете последних данных по перекрестной релаксации. Многие из опубликованных ранее результатов могут оказаться справедливыми лишь в ограниченном диапазоне условий. [10]
За образованием гиперцикла должна следовать стадия обособления от среды - компартментализация. При этом кодирующие единицы объединяются в единую замкнутую цепь, воспроизводящую себя. Эта стадия моделирует уже клеточный уровень. Надо сказать, что Эйген и Шустер, а также Эбелинг провели расчеты, основанные на теории гиперциклов, а опыты Шпигельмана с размножением фага PQ в пробирке послужили для ее количественной проверки, причем получился хороший результат. [11]
Исходя из отсутствия каких-либо мембран или полупроницаемых барьеров в этих искусственным путем полученных материалах, вопрос о компартментализации как необходимом условии проявления сложной релаксации нуждается в пересмотре. [12]
После нуклеации гиперцикл должен пространственно обособиться от среды. Компартментализация необходима для использования функциональных ветвей, возникших вследствие мутаций. Компартментализованная система может индивидуализироваться путем соединения своих кодирующих единиц в единую цепь и воспроизводить всю эту цепь. В такой цепи, которая при участии лигаз будет циклической, гены, происходящие от данной единицы / ь локализуются в соседних положениях. [13]
С другой стороны, тонкая регуляция биосинтетических и биоэнергетических процессов в живой клетке осуществляется на основе пространственного разделения функциональных частей клетки, ее органоидов. Внутриклеточные мембраны служат для компартментализации внутреннего содержимого клетки. [14]
Активность пероксидазы возрастает и старение тканей ускоряется с увеличением концентрации фторидов. Нарушение естественных реакций обмена веществ наряду с изменениями структуры органелл и компартментализации клетки в конце концов приводят к ее гибели. [15]
Страницы: 1 2