Комплементарность
Cтраница 1
Комплементарность достигается за счет изменения конформации активного центра фермента под действием субстрата, а также перевода субстрата в переходное состояние, индуцируемого функциональными группировками активного центра фермента. [1]
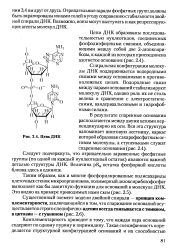 |
Цепь ДНК. [2] |
Комплементарность приводит к тому, что каждая пара оснований содержит по одному пурину и пиримидину. [3]
Комплементарность полинуклеотидных нитей РНК своим цистронам доказана методом гибридологического анализа. [4]
Комплементарность структуры активного центра фермента структуре субстрата лежит, следовательно, в основе как высокой активности, так и отчетливой субстратной специфичности ферментов. [5]
Если комплементарность между активным центром и переходным состоянием является существенной для ферментативного катализа, то нельзя исключить возможность синтеза искусственного фермента за счет конструирования подходящего активного центра. Один из возможных путей заключается в получении антитела к гаптеновой группе, которая адекватна переходному состоянию данной реакции. Связывающие центры таких антител должны быть комплементарны переходному состоянию и должны вызывать ускорения за счет переведения связанных субстратов в положение, похожее на переходное состояние. [6]
Если комплементарность между зондом и ДНК-мишенью отсутствует, то гибридизации не происходит, и мы получаем отрицательный результат. [7]
Доказательства комплементарности и антипараллельности двух цепей были получены методом определения частот ближайших соседей ( см. стр. Опыты, проведенные Джоссом и Корнбергом, убедительно свидетельствуют в пользу первой из этих двух альтернатив ( см. стр. [8]
Величина комплементарности нитей ДНК разного происхождения отражает степень генетического родства организмов, которым принадлежат исходные молекулы ДНК. Стопроцентная, или абсолютная, комоплементарность присуща нитям исходных молекул. [9]
ДНК-полимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( З - концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, Которая отщепляет не спаренный с матрицей нуклеотид затравки, После чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Му-Тации, нарушающие З - экзонуклеазную активность ДНК-полимера-Зы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной активности относительно полимеризующей, снижают темп мутирования генетического материала. [10]
ДНК-полимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( 3 -концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, Которая отщепляет не спаренный с матрицей нуклеотид затравки, после чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З - экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной актив - Поста относительно полимеризующей, снижают темп мутирования генетического материала. [11]
ДНК-пслимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( З - концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью которая отщепляет не спаренный с матрицей нуклеотид затравки, после чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З - экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной активности относительно полимеризующей, снижают темп мутирования генетического материала. [12]
Субституционалыюсть и комплементарность благ. [13]
Замечательное свойство комплементарности, благодаря которому между соответствующими основаниями возникают водородные связи, проявляется в том, что две соседние молекулы ДНК удерживаются друг около друга за счет содержащихся в них пар оснований Т - А и Ц - Г так, что получается двойная спираль. [14]
Это отношение комплементарности не имеет места, если мы ограничиваемся случаем 1-факторов. [15]
Страницы: 1 2 3 4