Константа - ингибирование
Cтраница 1
Константа ингибирования n - бензохинона в данном процессе равна 4 5 при 60 С. [1]
Здесь константа ингибирования / c 0 l и скорость зависит от первой степени интенсивности света. При этом предполагается, что атомы С1 исчезают при диффузии ( или конвекции) к стенкам по реакции первого порядка. Это более или менее хорошо согласуется с другими работами [30, 31] в этой области, хотя вследствие трудностей, возникающих при применении метода стационарных концентраций, все эти результаты должны быть приняты с некоторыми оговорками. Краггс [32], Алманд и Сквайр [32, 33] работали с очень низкими концентрациями Н2 и показали, что зависимость от интенсивности света изменяется от I a при низких концентрациях С12 ( - 0 01 мм рт. ст.) и низких интенсивностях света до 1Ц при больших концентрациях С12 ( - 450 мм рт. ст.) и больших интенсивностях света. [2]
Здесь константа ингибирования & г - 0 1 и скорость зависит от первой степени интенсивности света. При этом предполагается, что атомы G1 исчезают при диффузии ( или конвекции) к стенкам по реакции первого порядка. Это более или менее хорошо согласуется с другими работами [30, 31] в этой области, хотя вследствие трудностей, возникающих при применении метода стационарных концентраций, все эти результаты должны быть приняты с некоторыми оговорками. Краггс [32], Алманд и Сквайр [32, 33] работали с очень низкими концентрациями Н2 и показали, что зависимость от интенсивности света изменяется от I a при низких концентрациях С1г ( - 0 01 мм рт. ст.) и низких интенсивностях света до 1Цъ при больших концентрациях С12 ( - - 450 мм рт. ст.) и больших интенсивностях света. [3]
Сопоставление констант ингибирования / Ci для фенилфосфата и величин К для 2 4 - и 2 6-динитрофенилфосфатов показывает, что нитрогруппы способствуют связыванию, причем / г-нитрогруппа более эффективна в этом отношении, чем о-нитрогруппа. [4]
Величины констант ингибирования известны для различных процессов и ингибиторов. [5]
Эти вещества обладают низкой константой ингибирования связывания диазепама мебранами переднего мозга крыс. [6]
 |
Элюирование - нитрофенола на. [7] |
Определение констант связывания К и констант ингибирования / Ci для мицеллярно-каталитических реакций позволяет, по крайней мере качественно, интерпретировать влияние структуры субстрата на природу процесса комплексообразования с мицеллами и дает возможность сравнить константы связывания в мицеллярных системах с константами связывания в ферментативных реакциях. Ограниченное число данных такого рода не позволяет пока сделать каких бы то ни было широких обобщений. [8]
Отношение / Сю / к0бр определяется как константа ингибирования С пг. [9]
Полагая, что отношение величин К, для эфиров и амидов совпадает с отношением констант ингибирования ( Ki) для амида и изопропилового эфира N-ацетил-о - триптофана, равным 2 9, Бендер вычислил истинную величину / (, для этих эфиров. Вычисленные величины &3 для этих трех эфиров согласуются между собой, а величины &2 для четырех соединений хорошо коррелируют с порядком нук-леофильности, определенным на основании констант скоростей второго порядка щелочного гидролиза этих соединений. [10]
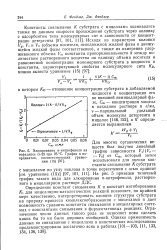
Из этого уравнения следует, что величина / 5о при постоянной концентрации субстрата однозначно определяет величину константы ингибирования. Заметим, что для частного случая весьма малых концентраций субстрата, когда AT [ S ] 1, значение / 50 численно равно величине, обратной константе ингибирования. [11]
Величину К [ E ] - I ] / [ EI ], которая представляет собой константу диссоциации комплекса фермента с ингибитором, называют константой ингибирования. Таким образом, при конкурентном ингиби-ровании увеличивается константа Михаэлиса, а максимальная скорость ферментативной реакции остается неизменной. [12]
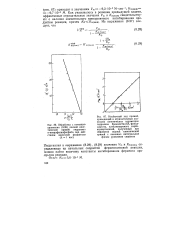
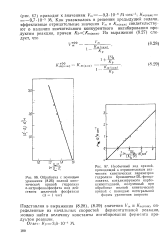
Подставляя в выражения (8.28), (8.29) значения Vm и / Ст ( каж), определенные из начальных скоростей ферментативной реакции, можно найти величину константы ингибирования фермента продуктом реакции. [14]
Страницы: 1 2 3