Константа - ингибирование
Cтраница 2
Подставляя в выражения (8.28), (8.29) значения Vm и / С Г1 ( каж), определенные из начальных скоростей ферментативной реакции, можно найти величину константы ингибирования фермента продуктом реакции. [16]
Зная, что обрыв преимущественно протекает вследствие взанмодействия свободных радикаяов с ингибитором, по концентрациям мономера н ингибитора ( 2 0 и 0 08 моль-л 1), начальиой скорости полимеризации ( 0 88 - 10 - 6 моль-л - - с 1) и константе ингибирования ( 1 2) вычислите начальные скорость инициирования и ялину кинетической цепи. [17]
Зная, что обрыв преимущественно протекает вследствие взаимодействия свободных радикалов с ингибитором, по концентрациям мономера и ингибитора ( 2 0 и 0 08 моль-л 1), начальной скорости полимеризации ( 0 88 10 - 6 моль л - 1 с -) и константе ингибирования ( 1 2) вычислите начальные скорость инициирования и длину кинетической цепи. [18]
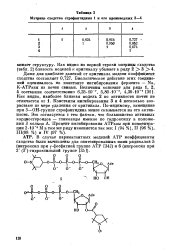 |
Матрица сходства строфантиднна 1 и его производных 2 - 4. [19] |
Константы ингибирования 3 и 4 несколько возрастают по мере удаления от оригинала. По-видимому, замещение при 3 - ОН-группе строфантиднна менее сказывается на его активности. Это согласуется с тем фактом, что большинство активных кардиост ероидов - гликозиды именно по гидроксилу в положении 3 кольца А. [20]
В таблице 3 приведены кинетические данные для гидролиза этилового эфира Кт-ацетил - Ь - тирозина, катализируемого а-хи-мотрипсином в присутствии конкурентного ингибитора, метилового эфира М - ацетил - В-фенилаланил - Ь - аланина. Определить значение константы ингибирования фермента DD-дипептидом, если начальная концентрация субстрата и начальное время реакции неизвестны. Определение концентрации непрореагировавшего субстрата в ходе ферментативной реакции проводилось в каждом случае через равные промежутки времени. [21]
 |
Ингибирование полимеризации винилацетата дурохино-ном. Инициатор - перекись бензо-ила. температура 45 С. [22] |
В табл. 3.7 приведены значения z для различных систем. Нетрудно заметить, что константы ингибирования, как и константы передачи цепи, сильно зависят от реакционной способности растущего радикала. [23]
Как правило, величина константы ингибирования для моносахари - ДОВ - ПРОДУКТОВ ферментативного гидролиза - близка к величине константы Михаэлиса для соответствующего субстрата. Аналоги или некоторые производные таких моносахаридов являются, по-видимому, наиболее эффективными конкурентными ингибиторами гликозидаз, известными к настоящему времени. [24]
Другим преимуществом является большая быстрота и расход очень небольших количеств фермента. Использование аффинной хроматографии для определения, например, констант ингибирования ферментов, по-видимому, весьма перспективно. Данные об объемах элюатов, содержащих фермент, на колонке с иммобилизованным ингибитором, полученные при использовании различных концентраций ингибитора в растворе, позволяют определить константы ингибирования как для связанного ингибитора, так и для ингибитора в растворе. Метод детально рассмотрен в гл. Большое преимущество этого метода состоит в том, что при использовании одного и того же ингибитора как для иммобилизации, так и для элюирования можно непосредственно сделать выводы о влиянии связей с носителем и природы носителя на изучаемое взаимодействие на основании совпадения или различий в определяемых величинах констант диссоциации. Следовательно, метод аффинной хроматографии открывает новые возможности не только для изучения взаимодействия биологически активных веществ, он перспективен также и для выяснения влияния микроокружения на образование этих комплексов. [25]
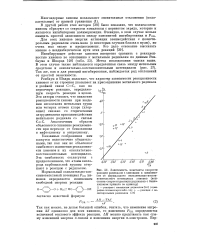
В другой работе этих авторов [38] было показано, что полигалоген-хиноны образуют со стиролом комплексы с переносом заряда, которые и являются ингибиторами полимеризации. Очевидно, в этом случае нельзя ожидать простой зависимости между константой ингибирования. [27]
Метод Диксона обладает тем преимуществом, что он позволяет определять значение константы ингибирования непосредственно из кинетических данных, не прибегая к дополнительным построениям. С другой стороны, вид графика в координатах Диксона не позволяет отличить смешанный тип ингибирования от конкурентного или неконкурентного ингибирования, как это обычно можно сделать при построении в координатах Лайнуивера-Берка. [28]
Лайнуивера-Берка, соответствующие различным концентрациям ингибитора, имеют вид семейства параллельных прямых. Предложить схему ингибирования ( приняв двухстадийный механизм ферментативной реакции) и рассчитать константу ингибирования. [29]
Точно так же - в разобранном выше случае с трансаминазой константы Михаэлиса и константы ингибирования продуктом реакции ( равновесные константы) совпадают, если субстратами являются аминокислоты, хотя различаются в случае кетокислот. [30]
Страницы: 1 2 3