Конъюгация - хромосома
Cтраница 2
Одним из критериев, позволяющим определять тип полиплоидии, является характер конъюгации хромосом в метафазе I деления мейоза. [16]
Самоопыляющиеся виды растений гомозиготны, а у гомозигот, как правило, наблюдается лишь обычная попарная конъюгация хромосом. Однако иногда у гибридов FI, полученных от скрещивания между двумя гомозиготными сортами, образуются одно или несколько типичных это бывает обусловлено тем, что родительские формы имеют различное строение хромосом. [17]
 |
Вес 1000 зерен ( в г диплоидных и тетраплоицных форм кукурузы. [18] |
Повышение плодовитости может также достигаться нарушением гомологии хромосом, т.е. переводом на бивалентный тип конъюгации хромосом. [19]
Основным методом, позволяющим определить различия в геномном составе у полиплоидов, является анализ конъюгации хромосом у межвидовых гибридов и полигаплоидов. [20]
Если сравнить формы из разных фенотипических групп, имеющие сходные цитологические особенности, то окажется, что само наличие определенного типа нарушений в конъюгации хромосом не обеспечивает однотипного изменения морфологических признаков растений. Например, присутствие мультивалентов характерно для FJ от скрещивания мутанта 204 с Л329, однако эта мутантная форма не отличается от сорта Л329 по высоте растений, длине колоса и озерненности. Мутант 206 / 2 также имеет тетравалент у гибридов FJ, Но по высоте он достоверно меньше, чем Лютесненс 329, не отличается от нее по озерненности, хотя имеет более длинный колос. [21]
В данном случае конъюгировавшие хромосомы взяты не из пахинемы мейоза, а из интерфазного ядра клетки слюнной железы личинки, в которой ( как это ни странно) происходит необычайно точная конъюгация хромосом и их участков. О свойствах хромосом слюнных желез говорится ниже ( стр. [22]
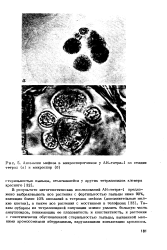 |
Аномалии мейоза в микроспорогенезе у АН-тетра-1 на стадии тетрад ( а и микроспор ( б. [23] |
Таким отбором из тетраплоидной популяции можно удалить большую часть анеуплоидов, понижающих ее плодовитость и константность, и растения с генотипически обусловленной стерильностью пыльцы, вызванной мелкими хромосомными аберрациями, нарушающими конъюгацию хромосом. [24]
Этот случай ясно показывает, что сходство между хромосомными комплексами соматических клеток не может служить доказательством сходства их структуры. Более надежные сведения может дать изучение конъюгации хромосом во время мейоза или ( у двукрылых) в клетках слюнных желез. Конъюгация часто бывает более или менее нарушена ввиду наличия структурных различий; в других же случаях она протекает нормально или почти нормально. В этих случаях конъ-югирующие хромосомы близки в структурном отношении или же совершенно гомологичны. Однако надо подчеркнуть, что даже совершенно нормальное образование бивалентов в первой метафазе мейоза не может служить доказательством полного структурного сходства хромосом скрещиваемых видов. [25]
Дарлингтон [46 ] описал 10 вероятных моделей квадривалентных ассоциаций хромосом и 10 ассоциаций других конфигураций. Так как у изучаемых амфидиплоицов и аутотетраплоидной кукурузы конъюгация хромосом, в основном, осуществляется квадри - валентами и бивалентами, нами был произведен учет только отдельных конфигураций квадривалентов и подсчет общего числа бивалентов без установления их типа. Частота каждого из типов ассоциаций представлена в табл. 9, а общий вид стадий диакинеза у изучаемых форм - на рисунке. [26]
Для того чтобы определить роль этих отклонений в возникновении различий в конъюгации хромосом у гибридов FJ и Лютесценс 329 были произведены два типа разбивки на классы. При первом типе разбивки мы объединили все клетки, содержащие только биваленты, независимо от числа терминальных хиазм, т.е. клетки, которые имели только закрытые, часть открытых и преждевременно разошедшихся бивалентов. Клетки с 20jj 2j составляли второй класс, с 19Ц 41 - третий, а клетки, содержащие мультивалентные конфигурации хромосом, - остальные классы. [27]
Отсутствие перекреста у самцов дрозофилы объясняется тем, что процесс конъюгации хромосом в мейозе у самцов и самок различен. У самок происходит настоящий обмен участками между конъюгировавшими хромосомами, тогда как у самцов конъюгация хромосом происходит иначе и не приводит к обмену участками. [28]
Основная причина стерильности гибридов FI - несовместимость геномов, хромосом и отдельных генов, следствием чего является нарушение конъюгации хромосом в мейозе. [29]
Данные Сваминатана и Хугаса [15], Маркса [6, 16] и Хокса [ 17] показали, что эти гибриды почти полностью стерильны и в мейоае формируют в среднем 24д 24j - Такой тип конъюгации хромосом свидетельствовал о четких различиях между геномами исходных родительских видов. [30]
Страницы: 1 2 3 4