Мембрана - клетка
Cтраница 2
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, и это придает поверхности мембран биологическую специфичность. [16]
Связывание ФВВ с мембранами клеток может сопровождаться структурными изменениями в липидах и белках мембран, приводящими к изменению проницаемости мембран, в частности, ФВВ. Важную роль при этом играют изменения текучести липидов, что может сказаться на изменеии биодоступности ЛВ. [17]
Сахаров и аминокислот через мембраны клеток почек и кишечника называется симпортом. [18]
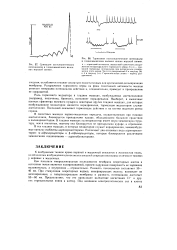 |
Суммация постсинаптических потенциалов в гладкомышечных волокнах морской свинки.| Тормозные постсинаптические потенциалы в гладкомышечном волокне кишки морской свинки. [19] |
Как показали микроэлектродные исследования мембрана секреторных клеток в состоянии покоя является поляризованной, причем наружная поверхность ее заряжена положительно, а внутренняя - отрицательно. Разность потенциалов составляет 30 - 40 мв. При стимуляции секреторных нервов, иннервирующих железу, возникает не деполяризация, а гиперполяризация мембраны и разность потенциалов достигает 50 - 60 мв. Предполагают, что это происходит вследствие нагнетания С1 - и других отрицательных ионов в клетку. [20]
Градиент ионной концентрации в мембране клетки вырабатывает внутри клетки электрический потенциал - 70 мВ относительно ее окружения. При этом потоки натрия и калия сразу направляются в обратную сторону; в течение миллисекунд внутренний потенциал клетки становится 50 мВ относительно внешнего окружения. Это изменение полярности быстро распространится через клетку, заставляя нервный импульс распространиться по всему аксону до его пресинаптических окончаний. Когда импульс достигнет окончания аксона, открываются управляемые напряжением кальциевые каналы. Это вызывает освобождение нейротрансмиттерных молекул в синаптическую щель и процесс распространяется на другие нейроны. [21]
На микрофотографиях обычно между мембранами соприкасающихся клеток виден просвет шириной от 100 до 200 А. По некоторым данным ионы кальция способствуют слипанию клеток. Возможно, что этот эффект связан с локальной поляризацией мембран. [22]
Витамин Е накапливается в мембранах клеток и действует как антиоксидант, прерывая цепи свободноради-кальных реакций. Это препятствует пероксидации полиненасыщенных жирных кислот фосфолипидов. При этом происходит перенос фенольного водорода на свободный радикал переокисляющейся ненасыщенной жирной кислоты. Продукт превращения токоферола, не содержащий свободного радикала, конъюгируется с глюкуроно-вой кислотой в печени и выводится с желчью. [23]
Различная скорость проникновения аминокислот через мембраны клеток, установленная при помощи метода меченых атомов, свидетельствует о существовании в организме активной транспортной системы, обеспечивающей перенос аминокислот как через внешнюю плазматическую мембрану, так и через систему внутриклеточных мембран. Несмотря на тщательные исследования, проведенные в разных лабораториях, тонкие механизмы функционирования активной системы транспорта аминокислот пока не расшифрованы. Очевидно, таких систем существует несколько. Майстером предложена оригинальная схема транспорта нейтральных аминокислот через плазматическую мембрану, которая, по-видимому, активна в почечных канальцах, слизистой оболочке кишечника и ряде других тканей. [24]
Структурные белки входят в состав мембран клеток и отличаются высокой степенью гидрофобности. Так называемые интегральные белки способствуют стабилизации клеточных мембран, но не обладают какой-либо функциональной активностью. К структурным белкам относятся также белки межклеточного матрикса, такие, как коллаген и ретикулин. Одним из основных компонентов связок является эластин, а кожи - коллаген. Что касается волос и ногтей, то они в основном состоят из очень прочного белка - кератина. [25]
Внутренняя и внешняя стороны протоплазматичесшх мембран клеток, по-видимому, имеют различный состав липидов. Изменение формы эритроцитов, вызываемое различными лекарственными треп аратами [86], возможно, объясняется именно различием их липидного состава, поскольку одно и то же вещество может вызвать различное расширение монослоев, состоящих из различных липидов. В этом случае бислой предполагается состоящим из двух как бы независимых моиослоев. [26]
Насосы активно перемещают ионы через мембрану клетки для поддержания градиентов концентрации. Каналы пропускают ионы выборочно и управляют их прохождением через мембрану. Некоторые каналы открываются или закрываются распространяющимся через мембрану электрическим потенциалом, тем самым обеспечивая быстрое и чувствительное средство изменения ионных градиентов. Другие типы каналов управляются химически, изменяя свою проницаемость при получении химических носителей. [27]
Мембраны клеток печени в отличие от мембраны клеток жировой ткани и мышечных волокон свободно проницаемы для глюкозы и в отсутствие инсулина. Предполагают, что этот гормон действует непосредственно на углеводный обмен печеночных клеток, активируя синтез гликогена. [28]
Гемолиз - механический или химический разрыв мембраны клетки с выходом содержимого во внеклеточную среду. [29]
Гормоны, сравнительно легко проникающие через мембрану клетки ( стероидные и в некоторой степени тиреоидные гормоны), оказывают непосредственное специфическое влияние на определенные внутриклеточные структуры. Их действие развертывается и осуществляется длительно, так как они, как правило, влияют на процессы транскрипции, осуществляющиеся в клеточном ядре, изменяя процессы синтеза определенных клеточных белков. [30]
Страницы: 1 2 3 4