Мембрана - эндоплазматический ретикулум
Cтраница 2
Эндоплазматический ретикулум - это внутриклеточная сеть взаимосвязанных, ограниченных мембраной трубочек и пузырьков. Мембрана эндоплазматического ретикулума образует единое целое с ядерной мембраной. [16]
Однако в эукариотах рибосомы после разрыва пептидного якоря еще обнаруживают существенное сродство к мембране эндоплазматического ретикулума. В опытах in vitro полная диссоциация рибосом от мембран эндоплазматического ретикулума достигается лишь путем совместной обработки микросом пуромицином и высокой ионной силой. [17]
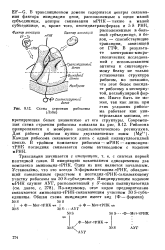 |
Схема строения рибосомы ( Лэйк. [18] |
Иначе быть не может, так как лишь при этих условиях рибосома работает как универсальная машина, се-кретирующая белок независимо от его структуры. Современная схема строения рибосомы показана на рис. 8.12. Рибосома прикрепляется к мембране эндоплазматического ретикулума. Каждая рибосома связывается лишь с одной полинуклеотидной депью. [19]
 |
Схема, демонстрирую - цепи. [20] |
Уже давно было показано что во всяком случае эукариотиче-ские рибосомы прикрепляются к мембране своей большой ( 60S) субчастицей. По-видимому, существует специальный участок на 60S субчастице, который имеет сродство к мембране эндоплазматического ретикулума, и, таким образом, все рибосомы прикрепляются к мембране строго определенной точкой, в одной и той же ориентации. [21]
Возможно существование каких-то регуляторных белков или малых рибонуклеопротеидов, которые взаимодействуют с транслирующей рибосомой и избирательно останавливают или затрудняют элонгацию в определенных местах. Известен пример таких специфичных репрессоров элонгации в эукариотах: это рибонуклеопротеид-ная частица, содержащая 7S РНК; частица узнает особую N-концевую гидрофобную последовательность образующегося полипептида на транслирующей рибосоме, присоединяется к рибосомам и останавливает элонгацию до тех пор, пока рибосома не вступит во взаимодействие с мембраной эндоплазматического ретикулума ( см. B. Не исключено, что подобные механизмы используются для регуляции скорости элонгации на других стадиях синтеза белка, например, на определенных стадиях сворачивания белка или сборки белка на транслирующей рибосоме. [22]
Помимо ядра и митохондрий в клетках эукариот существует и функционирует значительное число других органелл. Среди них следует в первую очередь упомянуть систему эндоплазматического ретикулума, которая представляет собой большое число ограниченных мембранами слоев, пузырьков, трубочек. Мембраны эндоплазматического ретикулума образуют, по-видимому, единую систему, связанную также с ядерной мембраной. Различают гладкий эндоплазматический ретикулум, имеющий трубчатое строение, в котором происходит в основном метаболизм липидов. Кроме того, широко представлен шероховатый эндоплазматический ретикулум, на внешних поверхностях которого закреплены рибосомы. На этих рибосомах происходит синтез многочисленных мембранных белков. [23]
Сведения по цитологии грибов связаны в основном с исследованием строения клетки дрожжевых и дрожжеподобных грибов. В дрожжевой клетке обнаружен эндоплазматический ретикулум ( ЭР), представляющий собой систему пузырьков или цистерн и канальцев, соединяющихся с нуклеолеммой и цитоплазмати-ческой мембраной. Мембраны эндоплазматического ретикулума обеспечивают продвижение различных веществ по грибной клетке, представляют собой активные поверхности для локализации ферментов, а следовательно, и метаболических процессов. Есть предположение об участии ЭР в синтезе липидов и углеводов. [24]
Посттрансляционная модификация белка включает следующие процессы: химическую модификацию белка ( часто отсутствует) - метилирование по аминогруппе лизина и аргинина, фос-форилирование по ОН-группе серина, окисление лизина, пролина и др.; связывание простетической группы; связывание между собой субъединиц олигомерного белка; частичный протеолиз. Например, ттосттрансляционная модификация при биосинтезе гликопротеинов происходит следующим образом. Полисомы связаны с внешней поверхностью мембраны эндоплазматического ретикулума клеток через большую субъединицу рибосомы. [25]
Кроме секретируемых белков, растущие полипептидные цепи ряда встроенных в мембрану белков также характеризуются временной сигнальной N-концевой последовательностью. Одним из первых изученных примеров такого рода был гликопротеид вируса везикулярного стоматита, который вместе с хозяйской мембраной участвует в построении вирусной оболочки. Этот белок, как оказалось, синтезируется с N-концевой сигнальной последовательностью, очень похожей на таковую секретируемых пребелков; сигнальная последовательность необходима для присоединения транслирующей рибосомы к мембране эндоплазматического ретикулума; дальнейший синтез белка идет, таким образом, на мембраносвязанных рибосомах; в ходе элонгации N-концевая последовательность из 16 аминокислотных остатков отщепляется в мембране. Другими словами, все это не отличимо от ситуации в случае водорастворимых секретируемых белков. [26]

Полярные липиды, к числу которых относятся только что рассмотренные фос-фоглицеролы, сфинголипиды и гликоли-пиды, не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях. Фосфоглицеролы, синтезируемые ферментами эндоплазматическо-го ретикулума, встраиваются в основном в липидный бислой ретикулума. Общая площадь эндоплазматического ретикулума особенно велика в клетках печени и поджелудочной железы. Мембраны эндоплазматического ретикулума служат предшественниками мембран аппарата Гольджи. Эти пузырьки часто сливаются с плазматической мембраной. Фосфоглицеролы могут переноситься из эндоплазматического ретикулума в митохондрии также при помощи транспортных белков. Таким образом, в клетке существует поток вновь синтезированных полярных липидов, направленный к различным типам клеточных мембран. [28]
Восстановлением дисульфидных связей и их последующим реокислением был получен проинсулин с - 70 % - ным выходом. На рис. 2 - 41 приведена первичная структура проинсулина свиньи, состоящего из 84 аминокислот. В процессе биосинтеза на первом этапе образуется фрагмент, содержащий в соответствии с так называемой сигнальной гипотезой ( разд. N-концевую сигнальную последовательность с информацией о том, что последующая последовательность проиисулина строится прямо на мембране эндоплазматического ретикулума и что этот инсулиновый предшественник уже в процессе синтеза должен уходить через мембрану по внутренним каналам ЭР. Еще во время биосинтеза сигнальный пептид отщепляется особой сигнальной пептидазой. Проинсулин затем идет в аппарат Гольджи, где расщепляется на инсулин и С-пелтид и в кристаллической форме в присутствии цинка накапливается в визикулах. При соответствующем физиологическом раздражении происходит экзоцитоз и содержимое визикул попадает в кровь. [29]
 |
Внешние факторы, влияющие на биотрансформацию ксенобиотиков. [30] |
Страницы: 1 2 3