Механизм - репликация
Cтраница 1
 |
Схема образования репликативной формы ДНК фага, содержащего с зрелом вирусе однонитевую ДНК. [1] |
Механизм репликации, используемый для производства плюс епей, также нетрадиционен. Процесс начинается с образования в определенной точке в составе RF-I плюс - - цепи гидролитического разрыва, приводящего к появлению свободной 3 -гид-роксигруппы и, соответственно - концевой фосфомоноэфирной группы. У бактериофага ipX 174, механизм репликации которого изучен наиболее полно, разрыв происходит под действием специального, программируемого фаговой ДНК белка, известного как белок А. Разрывы, подобные тому, который образуется при действии на репликативную ДНК белка А, часто фигурируют в литературе под названием ник ( от англ. Двунитевая кольцевая ДНК с ником в плюс-цепи называется репликативной формой II или RF-II. Появившаяся гидро-ксигруппа играет роль праймера, с которого начинается процесс элонгации. Такая элонгация по своей сути означает синтез новой плюс-цепи, ковалентно связанной с исходной. [2]
Механизм репликации одноцепочечной вирусной РНК неизвестен. Так как цепь нуклеиновой кислоты характеризуется определенным направлением, цепь, комплементарная к исходной, должна определять совсем другой белок, если только последовательность оснований РНК не является совсем уж особенной. [3]
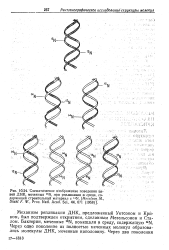 |
Схематическое изображение поведения це -. [4] |
Механизм репликации ДНК, предложенный Уотсоном и Криком, был подтвержден открытием, сделанным Мезельсоном и Ста-лом. Через одно поколение из полностью меченных молекул образовались молекулы ДНК, меченные наполовину. [5]
Механизм асинхронной репликации данных применяют в РБД, в которых фрагменты ( ЛБД) связаны между собой ненадежными или имеющими низкую пропускную способность каналами связи. В таких РБД одновременная корректировка данных может привести к невозможности завершения распределенной транзакции. В дальнейшем рассматривается первая дисциплина выполнения транзакций. [6]
Для выяснения механизма репликации бактериальной хромосомы незаменимую роль сыграл анализ разнообразных мутантов, нарушающих репликацию ДНК - Синтез ДНК - функция жизненно важная, и мутации, инактивирующие ферменты синтеза ДНК, леталь-ны. [7]
Модель Кэрнса объясняет механизм репликации кольцевой ДНК - Согласно этой модели удвоение всегда начинается с одной и той же точки и идет всегда в одном направлении с помощью своеобразного шарнирного механизма; шарнир расположен в стартовой точке и дает возможность свободно вращаться неудвоенной части молекулы. Первоначально модель Кэрнса основывалась на радиоавтографических данных, свидетельствовавших о существовании репликационнои вилки; Шарнир может возникнуть в результате поочередного действия эндонуклеазы и лигазы, разрывающих и вновь соединяющих кольцевую молекулу ДНК - Радиоавтографические данные Кэрнса можно объяснить двумя вилками, которые движутся от общего начала в противоположном направлении. Такой двунаправленный синтез ДНК обнаружен в клетках кишечной палочки и подтверждается методами генетического анализа. Несмотря на то, что проблема раскручивания ДНК продолжает оставаться дискуссионной, все же результаты исследований указывают на то, что каждая из-исходных цепей ДНК полностью сохраняется в течение всего периода репликации. [8]
Описанные в настоящем разделе механизмы репликации справедливы по крайней мере для других бактерий. [9]
В учебнике рассмотрены принципы структурной организации ДНК и РНК механизмы репликации, репарации и рекомбинации ДНК, описаны механизмы транскрипции, посттранскрипционной модификации РНК и сплайсинга, особенности репликации н транскрипции геномов ДНК - и РНК-содержащих вирусов. [10]
Как будет видно из дальнейшего, особое значение для механизмов репликации линейных молекул ДНК имеет структура их концевых участков. У линейных ДНК-геномов не бывает невыразительных концов. Соответствующие участки ( рис. 134) могут иметь прямые концевые повторы длиной от сотни и более ( например, ДНК фага Т7) до тысяч ( Т - четные фаги и др.) пар нуклеотидов. При этом если у фага Т7 все геномные молекулы ДНК идентичны, то молекулы ДНК Т - четных фагов существенно различны, даже когда они образованы в одной клетке, зараженной единственной фаговой частицей; геномы Т - четных фагов ( и ряда других вирусов) характеризуются так называемыми кольцевыми перестановками. [11]
 |
Концевые участки вирусных линейных двухнитевых ДНК-геномов. [12] |
Как будет видно из дальнейшего, особое значение для механизмов репликации линейных молекул ДНК имеет структура их концевых участков. У линейных ДНК-геномов не бывает невыразительных концов. Соответствующие участки ( рис. 134) могут иметь прямые концевые повторы длиной от сотни и более ( например, ДНК фага Т7) до тысяч ( Т - четные фаги и др.) пар нуклеотидов. При этом если у фага Т7 все геномные молекулы ДНК идентичны, то молекулы ДНК Т - четных фагов существенно различны, даже Когда они образованы в одной клетке, зараженной единственной фаговой частицей; геномы Т - четных фагов ( и ряда других вирусов) характеризуются так называемыми кольцевыми перестановками. [13]
В учебнике рассмотрены принципы структурной организации ДНК и РНК, механизмы репликации, репарации и рекомбинации ДНК, описаны механизмы транскрипции, посттранскрипционной модификации РНК и сплайсинга, особенности репликации и транскрипции геномов ДНК - и РНК-содержащих вирусов. [14]

Страницы: 1 2 3 4