Механизм - репликация
Cтраница 2
Результаты этих опытов не совместимы ни с консервативным, ни с дисперсивным механизмом репликации ДНК; они согласуются с представлением о полуконсервативном механизме. При репликации цепи раскручиваются и расходятся. На каждой одиночной цепи происходит синтез новой комплементарной цепи. В этом процессе участвуют ДНК-по-лимеразы. Функция их проста: они связывают между собой нуклеотиды, расположившиеся путем спаривания оснований в правильном порядке, и таким образом синтезируют новую полинуклеотидную цепь. Биохимические механизмы показаны на рис. 2.15 и поясняются в подписи к этому рисунку. [16]
Полуконсервативная модель репликации ДНК - общепринятая в настоящее время модель, объясняющая механизм репликации ДНК. Этот механизм очень ярко иллюстрируют классические исследования Меселсона и Сталя. [17]
Мы уже видели и найдем тому много подтверждений дальше, что в клетке существует механизм репликации РНК на ДНК. Однако валовый состав РНК не соответствует валовому составу ДНК. [18]
Можно отметить, что наиболее широко представлены в литературе данные по воспроизведению генома клетки и биогенезу ядер; значительно меньше рассмотрены механизмы репликации других структур и буквально единичные сообщения относятся к вопросам синтеза капсул. В целом можно отметить, что многие органеллы в клетке достаточно автономны ( например, митохондрии и хромопласты) и способны сами синтезировать необходимые компоненты и структуры. В других случаях синтез проходит через стадии предшественников. Имеется вполне достоверное предположение, что рибосомный белок существует в микробной клетке в виде определенного фонда или запаса, откуда затем он включается в эосомы и неосомы, которые в свою очередь являются исходными структурами при построении рибосом. [19]
 |
Фенотипическая изменчивость роста у человека. Все эти дети одного возраста. [20] |
Необходимо ясно представлять себе, что взаимодействие между дискретной и непрерывной изменчивостью и средой делает невозможным существование двух организмов с идентичными фенотипами. Механизм репликации ДНК при митозе столь близок к совершенству, что возможности генетической изменчивости у организмов с бесполым размножением очень малы. Поэтому любая видимая изменчивость у таких организмов почти наверное обусловлена воздействиями внешней среды. Что же касается организмов, размножающихся половым путем, то у них есть широкие возможности для формирования генетической изменчивости. Практически неограниченными источниками генетической изменчивости служат кроссинговер и независимое расхождение хромосом во время мейоза, а также слияние гамет во время оплодотворения. Вкратце эти процессы описаны ниже. [21]
Генетический материал любой клетки представлен ДНК, информационные свойства которой определяются специфической последовательностью четырех нуклеотидов в полинуклеотидной цепи. Полуконсервативный механизм репликации ДНК, в результате которого из одной родительской двухцепочной молекулы образуются две дочерние молекулы, содержащие по одной родительской и одной вновь синтезированной комплементарной полинуклеотидной цепи, наилучшим образом обеспечивает идентичность исходной и синтезированных молекул и, следовательно, сохранность видоспецифической наследственной информации в ряду поколений клеток и организмов ( см. гл. [22]
Поскольку механизм репликации до конца еще не ясен, образование катенановых олигомеров можно представить с помощью двух механистических моделей. Согласно первой модели, которая объясняет также образование димеров, молекулы кольцевой двунитчатой ДНК сначала спариваются, затем разрываются и вновь реком-бинируют. [23]
Полимеризация дочерней ДНК на матрице ДНК приводит к ее удвоению или репликации. Для реализации механизма репликации необходима матрица - расплетенная цепь ДНК, субстраты, участвующие в полимеризации ДНК, ферменты, катализирующие этот процесс, ионы Mg2, а также белковые факторы, обеспечивающие деспирализацию двухнитевой ДНК. У прокариот ДНК имеет форму кольца, причем в определенном ori - сайте origin - начало репликации) цепи расходятся и образуются две репликативных вилки, движущиеся в противоположных направлениях. У эукариот имеется большое число ori - сайтов, и репликация проходит одновременно на многих участках ДНК. В точках начала репликации отмечено большое количество АТ пар оснований, соединенных всего лишь двумя водородными связями, что способствует более легкому разрыву и расхождению цепей. [24]
Описанные в настоящем разделе механизмы репликации справедливы по крайней мере для других бактерий. [25]
В белках л-электронные системы сравнительно слабо проявляют себя. Исключительного развития эти системы достигают в соединениях, составляющих механизмы репликации и передачи наследственных признаков. Общей чертой биологически активных структур является сочетание в них областей ( групп атомов), богатых энергией, групп, содержащих объединенные и обширные я-орбитали, и участков, разделяющих те и другие. [26]
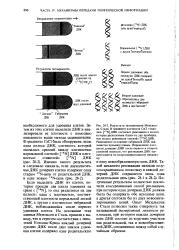
Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсона и Сталя позволил также отвергнуть так называемый дисперсивный механизм репликации, при котором каждая дочерняя цень ДНК состоит из коротких участков как родительской, так и новообразованной ДНК, соединенных между собой случайным образом. [28]
Механизм репликации, используемый для производства плюс епей, также нетрадиционен. Процесс начинается с образования в определенной точке в составе RF-I плюс - - цепи гидролитического разрыва, приводящего к появлению свободной 3 -гид-роксигруппы и, соответственно - концевой фосфомоноэфирной группы. У бактериофага ipX 174, механизм репликации которого изучен наиболее полно, разрыв происходит под действием специального, программируемого фаговой ДНК белка, известного как белок А. Разрывы, подобные тому, который образуется при действии на репликативную ДНК белка А, часто фигурируют в литературе под названием ник ( от англ. Двунитевая кольцевая ДНК с ником в плюс-цепи называется репликативной формой II или RF-II. Появившаяся гидро-ксигруппа играет роль праймера, с которого начинается процесс элонгации. Такая элонгация по своей сути означает синтез новой плюс-цепи, ковалентно связанной с исходной. [29]
Все эти различия становятся легко объяснимыми, если принять, что переход от ранней к поздней стадии репликации ДНК фага Я связан с внесением более или менее случайных разрывов в разные цепи 9-молекулы. Появление при этом молекулы а-типа становится понятным из рис. 143; понятно также, почему при этом не появляются новые потребности в ферментативном обеспечении: на ранней и поздней стадиях функционируют, по существу, одни и те же репликационные вилки. Все это дает основание отличать механизм репликации ДНК фага Я от схемы классического разматывающегося рулона. [30]
Страницы: 1 2 3 4