Механизм - сорбция
Cтраница 3
В зависимости от механизма сорбции - молекулярная и хемосорбц ионная хроматографии. [31]
В зависимости от механизма сорбции - молекулярная и хемосорбционная хроматографии. [32]
В зависимости от механизма сорбции - молекулярная и х е м о с о р б ц и о н п а я х рома т о г р а ф и и. [33]
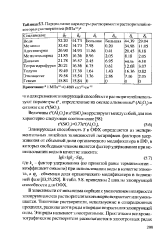 |
Парциальные параметры растворимости растворителей некоторых растворителей ( МПа1С. [34] |
В зависимости от механизма сорбции с увеличением полярности элюирующая сила растворителя закономерно возрастает или уменьшается. Типичные растворители, используемые в сепарационных процессах, располагают в ряды в порядке возрастания элюирующей силы. Эти ряды называют элюотропными. [35]
Принимая в качестве определяющего механизма сорбции ограниченную молекулярную растворимость воды в целлюлозе, следует иметь в виду, что для систем, в которых образуются интенсивные водородные связи ( в данном случае между молекулами воды и гидроксиль-ными группами целлюлозы) часто наблюдается своеобразная температурная зависимость этой растворимости. При повышении температуры водородные связи разрушаются, и совместимость должна уменьшаться. Действительно, система целлюлоза - вода обнаруживает признаки систем с нижней критической температурой совместимости. При понижении температуры совместимость ( величина сорбции, степень набухания целлюлозы в воде) увеличивается, а при повышении температуры - уменьшается. Область распада водородных связей, выше которой начинает сказываться увеличение совместимости за счет повышения кинетической энергии компонентов, лежит приблизительно при 50 - 70 С. [36]
К дифференциации по механизмам сорбции разделяемых веществ следует относиться с пониманием ее схематичности. [37]
Распространенным представлением о механизме сорбции паров является представление о конденсации паров в ультрапорах сорбентов. [38]
При использовании хелатных сорбентов механизм сорбции обусловлен комплексообразованием, а применение анионитов для концентрирования примесей металлов было бы невозможным бе образования последними анионных комплексов. При простом ионном обмене обычно удобнее перевести элемент матрицы в отличную от примесей ионную форму. В работе [61] разработана методика определения большого числа примесей в соединениях титана, тантала и ниобия во фторидной системе. В этой системе элементы матрицы находятся в анионной форме в виде комплексов MF6 ( 6 - rt) -, в то время как примеси остаются в виде катионов и могут быть сорбированы катионитом. На примере этой системы видно преимущество ионообменного концентрирования, позволяющего выделить примеси в динамических условиях при невысоких коэффициентах распределения. [39]
С целью детального изучения механизма сорбции и структуры сорбированной воды были привлечены методы ядерного магнитного резонанса ( ЯМР) и диэлектрический. Методом спин-эхо было показано, что зависимость спин-спиновой релаксации Т2 от влагосодержания подобна изотерме сорбции ( десорбции) и имеет точки перегиба, соответствующие границам физико-химической, осмотической и капиллярной влаги в торфе. [40]
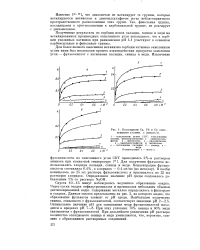 |
Поглощение Са, РЬ и Си окисленными углями, а ( ммоль / г. [41] |
Для более полного выяснения механизма сорбции катионов окисленным углем нами был исследован процесс взаимодействия продуктов окисления угля - фульвокислот с катионами кальция, свинца и меди. [42]
Если связывание идет по механизму обратимой сорбции с мгновенным установлением равновесия между твердой и жидкой фазами вида Ст KdC, то это учитывают в уравнениях ( 61) и ( 75) заменой v и D на отношения v / R и D / R, где Я - фактор задержки, зависящий от коэффициента распределения, пористости и плотности твердой фазы. [43]
Ограниченность наших представлений о механизме сорбции кислых белков на оксиапатите с еще большим основанием следует отнести к механизму сорбции основных белков. Вероятно, в ней участвуют кулоновские взаимодействия положительно заряженных остатков аминокислот на поверхности белка с отрицательно заряженными остатками фосфорной кислоты, которые тоже должны быть на поверхности оксиапатита. Об этом говорит более прочная сорбция полилизина и полиаргинина, чем других, более нейтральных полипептидов. Но вполне возможно, что и иные виды взаимодействия здесь играют важную роль. [44]
Одна из гипотез о механизме сорбции макромолекул ГМЦ на поверхности волокон целлюлозы [315, 481, 482, 680, 770] предполагает, что при соприкосновении с раствором ГМЦ начинается быстрая адсорбция цепей ГМЦ на открытой поверхности целлю-люзы, причем гибкие макромолекулы адсорбируются через сегменты или короткие последовательности цепей. Между адсорбированными сегментами существуют неадсорбированные участки, образующие мостики, или иетли, простирающиеся в раствор. В раствор могут простираться и неадсорбированные сегменты у концов цепей. Предполагается, что характер адсорбционных сил определяется образованием водородных свзей между доступными гидроксилами. После заполнения в основном свободной поверхности целлюлозы сорбированными цепями ГМЦ начинается более медленная стадия сорбции макромолекул ГМЦ на уже сорбированные ГМЦ. [45]
Страницы: 1 2 3 4