Миоглобин
Cтраница 4
Вследствие этого миоглобин насыщается кислородом уже при низком парциальном давлении, таким образом при сокращении мышц может быть компенсирован недостаточный приток кислорода из кровеносной системы. [46]
Гемоглобин и миоглобин - комплексы железопорфиринов с белками, выполняющие функцию фиксации и транспорта молекулярного кислорода в организмах животных. Цитохромы, имеющие аналогичную принципиальную структуру, выполняющие роль переносчика электрона в схемах фотосинтеза, дыхания, окислительного фос-форилирования и др. окислительно-восстановительных реакциях, найдены у всех животных, растений и микроорганизмов. Хлорофиллы - главные участники процессов фотосинтеза - содержатся в высших растениях, водорослях и фотосинтезирующих бактериях. [47]
Гемоглобины и миоглобины образуют группу белков, которые лучше всего подходят для исследования влияния белков на константу равновесия в процессе связывания одного лиганда, а именно кислорода. Все эти белки содержат один и тот же железопор-фириновый комплекс и, за исключением некоторых весьма редких мутантов, один и тот же аксиальный лиганд. Кроме того, все белки, о которых идет речь, обладают весьма сходной третичной структурой. Тем не менее величина константы равновесия связывания кислорода, а также гомотропное и гетеротропное взаимодействия для них изменяются в широких пределах. Начиная с новаторской ра-боты Кендрью и Перутца с сотрудниками по миоглобину кашалота и гемоглобинам человека и лошади, наиболее детальные сведения о структуре ряда гемоглобинов и миоглобинов получены методом рентгеноструктурного анализа. Благодаря тому интересу, который представляют для медицины мутантные белки, за последние годы многие мутантные формы гемоглобина были выделены и изучены, так что можно исследовать влияние замены даже одной аминокислоты на структуру белка и его сродство к кислороду. [48]
Гемоглобин и миоглобин в своих ферри-формах обнаруживают очень слабую пероксидазную и каталазную активность, однако ферро-пероксидаза обратимо с молекулярным кислородом не соединяется. Ионизированное железо также катализирует разложение перекиси водорода, но не соединяется с кислородом. [49]
Гемоглобин и миоглобин, пероксидаза и каталаза соединяются с очень большим числом различных веществ путем простых бимолекулярных реакций, включающих замещение молекулы воды или группы ОН. [50]
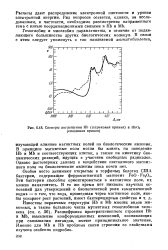 |
Спектры поглощения НЬ ( штриховая кривая и НЬО8 ( сплошная кривая. [51] |
Гемоглобин и миоглобин парамагнитны, в отличие от подавляющего большинства других биологических молекул. [52]
Гемоглобин и миоглобин - белки, содержащие соединение переходного металла, гем, в качестве простетической группы. Изучение такого рода веществ относится к области бионеорганической химии, о которой уже упоминалось выше ( с. Здесь уместно рассказать о проблемах бионеорганической химии в связи с биофизикой. [53]
Все точки миоглобина ( рис. 26а) попадают в области низких энергий, и, следовательно, обсуждение их не представляет интереса. В лизоциме, как было указано в работе [91], 18 остатков оказываются в неблагоприятных областях. [54]
Вторичные структуры миоглобина и гемоглобина представлены главным образом а-спиралями. Восемь отрезков полипептидной цепи являются относительно прямолинейными а-спиралями; эти отрезки содержат 80 % аминокислотных остатков всей цепи, остальные 20 % остатков приходятся на места сгибов, соединяющие между собой - спиральные отрезки. При укладывании цепи ( третичная структура) образуется полость, в которой расположена гем-группа. [55]
Структура молекулы миоглобина установлена Кендрю с сотрудниками [175] в 1957 г. на основе трудоемких рентгено-структурных исследований. [56]
Остов молекулы миоглобина построен из восьми относительно прямолинейных сегментов, чередующихся с изогнутыми участками полипептидной цепи. Оказалось, что все ос-спи-ральные сегменты представляют собой правую а-спираль. На долю а-спи-ральных участков в молекуле миоглобина приходится почти 80 % всех аминокислотных остатков. Рентгенострук-турный анализ позволил также установить точное расположение каждой из R-групп, выступающих из изображенных на рисунке контуров и занимающих практически все свободное пространство между изогнутыми петлями молекулы. [57]
О принадлежности миоглобина и цепей гемоглобина к одному семейству белков свидетельствует также сравнение аминокислотных последовательностей миоглобина кашалота и а - и р-цепей гемоглобина лошади. Как показано на рис. 8 - 11, во всех трех цепях имеются 27 эквивалентных положений, в которых находятся идентичные аминокислотные остатки; кроме того, в других 40 положениях обнаруживаются близкие по своим свойствам остатки аминокислот, например аспарагиновая и глутаминовая кислоты, или изолейцин и валин. Таким образом, и здесь мы видим, что в аминокислотных последовательностях гомологичных белков имеется ряд инвариантных аминокислотных остатков и что для гомологичных белков характерно сходство их трехмерной структуры. [58]
Кривая для миоглобина на рис. 7.5 рассчитана по этому уравнению. [59]
Страницы: 1 2 3 4