Молекула - каротиноид
Cтраница 2
В световой фазе участвуют два ансамбля светопоглощающих пигментов - фотосистемы I и II. Функциональная единица фотосистемы I содержит около 200 молекул хлорофилла а и 50 молекул каротиноидов. Эти пигменты являются сборщиками энергии фотонов. В этом ансамбле имеется одна молекула пигмента Р700, играющего роль реакционного центра. Фотосистема II включает 200 молекул хлорофилла а, 200 молекул хлорофилла Ь, фи-кобилины или ксантофиллы. В темноте ( в основном состоянии) окислительно-восстановительный потенциал ( ОВП) Р700 равен 0 40 В. Вероятность отдачи электрона очень мала. При поглощении энергии экситонов Р700 переходит в возбужденное состояние, что сопровождается изменением ОВП до - 0 60 В. Это обеспечивает способность Р700 отдать электрон на молекулу НАДФ с величиной ОВП, равной - 0 32 В. Передача электрона осуществляется через посредников: Р700 - связанный ферре-доксин - растворимый ферредоксин - НДЦФ-редуктаза - НАДФ. Два электрона транспортируются двумя молекулами ферредоксина. Потеря молекулой Р700 электрона приводит к переходу его в окисленное состояние; возникает электронная дырка, подлежащая заполнению. В молекуле Р700 оно осуществляется второй цепью переносчиков электронов, связывающей обе фотосистемы. При освещении фотосистемы II экситоны поглощаются пигментом Р680, который переходит в возбужденное состояние. [16]
Однако при промывании спиртом первоначальная окраска восстанавливается и каротиноид выделяется в неизмененном виде. Менье и Винэ высказали предположение, что окраска появляется вследствие резонанса полярных структур в результате перехода электронов от молекулы каротиноида к адсорбенту. Примерами необратимых изменений могут служить превращение витамина А в ангидровитамин А, перегруппировка системы двойных связей в витамине D2, потеря биологической активности витамином Ki при адсорбции на окиси алюминия или окиси магния ( но не па сахаре), а также каталитическое окисление витамина С на животном угле. При хроматографировании раствора антрацена в петролейном эфире на колонке из окиси алюминия, проявлении петролейным эфиром и экспозиции хроматограммы на свету из колонки выделяется диантрацен. При адсорбции нафтацена на окиси алюминия из бензольного раствора и проявлении тем же растворителем адсорбент становится желтым, а часть его, обращенная к прямому дневному свету, становится оранжевой вследствие образования хнпона. Продукты присоединения тринитробензола к некоторым ароматическим углеводородам, терпенам и стероидам, а также соответствующие пикраты расщепляются при прохождении через колонку из окиси алюминия. [17]
Расположение молекул хлорофилла и каротиноидов в том виде, как его представляют себе Уолкен и Швертц, показано на фиг. Согласно этой схеме, площадь, приходящаяся на одну молекулу хлорофилла, составляет в среднем 2 7 нм2, если молекулы плотно упакованы, причем остается достаточно места для молекул каротиноидов. Опыты in vitro с монослоями хлорофилла на различных жидкостях показали, что порфириновые кольца могут быть наклонены под углом около 45, как это представлено на фиг. По мнению Гедхира [121, 122], молекулы каротиноидов располагаются, вероятно, параллельно ламеллам, а не под прямым углом к порфириновым кольцам молекул хлорофилла, как показано на фиг. Гедхир и Фрей-Висслинг [109] предполагают, что эти кольца образуют монослои на изогнутых поверхностях макромолекул липопротеида. [18]
СС кароонильной с0 и др. Большую роль в поглощении света пигментами растений играют хромофоры с этиленовой группировкой. Центральная часть молекулы каротиноидов представляет сооой длинную сопряженную цепь этиленовых связей. Единичный изолированный хромофор этилен оесцветен. [19]
Каротиноидами называют встречающиеся в клетках растений и микроорганизмов желто-оранжевые пигменты, часть которых биологически активна. По химическому строению это полиены, молекулы которых содержат до 40 атомов углерода. Если в молекуле каротиноидов содержатся только углерод и водород, то их называют каротинами, а если в молекуле содержится и кислород - то их называют ксантофилами. [20]
В то время как основная масса фотосинтетических пигментов способна только поглощать энергию света и передавать ее соседним молекулам, небольшая часть молекул хлорофилла участвует в осуществлении фотохимической реакции, т.е. преобразовании электромагнитной энергии в химическую. Процесс этот происходит в реакционных центрах, состоящих из первичного донора электронов, первичного акцептора и одного или более вторичных акцепторов электронов. Кроме того, в составе реакционных центров обнаружены молекулы каротиноидов и полипептидов. [21]
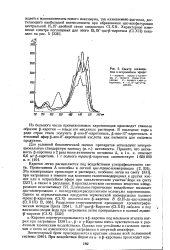 |
Спектр поглощения в петролейном эфире. [22] |
Каротин легко расщепляется под воздействием ультрафиолетового света. Эта изомеризация происходит в растворах, особенно легко на свету [331], при нагревании в темноте под влиянием галогеноводородных и других кислот или в петролейном эфире при каталитическом участии йода на свету 1337 ], а также в расплавах. Длительное термическое воздействие вызывает tywc - изомеризацию с последующим расщеплением молекулы каротиноидов. [23]
Однако некоторые каротиноиды состоят меньше чем из 8 изопреновых остатков ( например, кроцетин) или содержат большее число углеродных атомов, чем в изопреновых остатках ( например, азафрин; фиг. Для всех каротиноидов характерно, что в одном участке молекулы изопреновые остатки соединяются друг с другом в разрез с частным изопреновым правилом. За некоторыми исключениями ( например, азафрин) в центре молекулы каротиноидов два изопреновых остатка соединены друг с другом хвост в хвост. Это обстоятельство позволяет предполагать, что каротиноиды образуются при соединении двух идентичных дитерпенов. [24]
ПРОВИТАМИНЫ - соединения, являющиеся предшественниками витаминов, способные в организме превращаться в последние. Известны две группы таких соединений - карогпиноиды и стерины. Из каро-тннопдов путем окислительного расщепления образуется витамин А ( см. Ретинол) при условии, если хотя бы одна половина молекулы каротиноида соответствует строению витамина. Ряд стеринов является предшественниками витаминов О. Процесс превращения стеринов в витамины происходит под воздействием УФ-лучой и, в частности, имеет место в коже животных; этим приемом пользуются и при заводском изготовлении витаминных препаратов. [25]
В состав хлоропластов входят также каротиноиды и фико-эритрины. Каротины и их окисленная форма - ксантофиллы представляют собой систему сопряженных двойных связей. В хлоропласте они находятся между фитольными хвостами хлорофилла в виде пачек высотой 20 - 25 А, причем на молекулу хлорофилла приходится до трех молекул каротиноидов. Фико-эритрины и их окисленная форма - фикоцианины находятся в белковом слое и соотношение окисленной и восстановленной форм для всех трех видов пигментов определяется окислительно-восстановительным потенциалом среды. Липиды и белки составляют 35 - 50 % от сухого веса хлоропластов. В очень малом количестве ( 1 молекула на 300 - 400 молекул хлорофилла) содержатся ге-мопротеиды, цитохром f, цитохром Ь6, а также хиноны. Это свидетельствует о наличии цепочки передачи электронов. При возбуждении хлорофилл может обратимо отдавать или присоединять электрон. [26]
Производные каротиноидов, содержащие 20 атомов углерода в молекуле, играют важную роль в животном организме как витамины А. В настоящее время известно, что все светочувствительные пигменты, участвующие в процессе зрения животного организма, образуются путем соединения альдегидов витамина А, так называемых ретиненов, с протеинами. Способность каротиноида взаимодействовать с протеинами зависит от его формы; изменение в форме может изменить или устранить взаимодействие, причем как само взаимодействие, так и его прекращение направляется светом. Молекулы каротиноидов в отличие от других пигментов могут изменять свою форму путем перемены геометрической конфигурации своих двойных связей. Эти изменения прямо индуцируются действием поглощенного света, что приводит к образованию и выцветанию зрительных пигментов. Вполне возможно, что каро-тиноиды участвуют во всех типах светочувствительности живого мира. Примером является процесс зрения. [27]
Расположение молекул хлорофилла и каротиноидов в том виде, как его представляют себе Уолкен и Швертц, показано на фиг. Согласно этой схеме, площадь, приходящаяся на одну молекулу хлорофилла, составляет в среднем 2 7 нм2, если молекулы плотно упакованы, причем остается достаточно места для молекул каротиноидов. Опыты in vitro с монослоями хлорофилла на различных жидкостях показали, что порфириновые кольца могут быть наклонены под углом около 45, как это представлено на фиг. По мнению Гедхира [121, 122], молекулы каротиноидов располагаются, вероятно, параллельно ламеллам, а не под прямым углом к порфириновым кольцам молекул хлорофилла, как показано на фиг. Гедхир и Фрей-Висслинг [109] предполагают, что эти кольца образуют монослои на изогнутых поверхностях макромолекул липопротеида. [28]
 |
Схема тилакоида по Кальвину. [29] |
По строению ламеллы гран сходны с мембранами оболочки и стромы. Различие состоит главным образом в том, что в ламеллах гран один из слоев липидов в бимолекулярном слое замещен мономолекулярным слоем пигментов. Кроме того, в ламеллах тилакоид обнаружены регулярно повторяющиеся субъединицы - квантосомы, имеющие форму сплющенной сферы диаметром 180X160 А и толщиной 100 А. Каждая такая частица, помимо белка и липидов, содержит 230 молекул хлорофилла ( 160 - хлорофилла а и 70 - хлорофилла Ь), 48 молекул каротиноидов, цитохромы, пластохиноны, ферредоксин, марганец, медь. [30]
Страницы: 1 2 3