Молекула - фермент
Cтраница 1
Молекула фермента может состоять только из белка или из белковой и небелковой частей. Последняя получила название кофактора или простетической группы. Белковая часть молекулы фермента может быть построена из одной или нескольких полипептидных цепей, образующих сложные комплексы. Кофакторы имеют небольшую молекулярную массу и являются активной группой фермента. Ими могут быть производные витаминов, нуклеотидов или ионы металлов. Одни и те же кофакторы могут быть прочно связаны с белком или образовывать легко диссоциирующие комплексы. Одни и те же кофакторы могут входить в состав молекул разных ферментов. [1]
Молекулы ферментов могут узнавать форму частицы, несущей информацию, иными словами, они распознают информацию геометрического ( стерического) характера. В случае регулируемых ферментов она дает инструкции, согласно которым фермент либо катализирует, либо не катализирует определенную реакцию. Выяснилось, что в очень многих случаях молекула вещества-сигнала, несущая информацию, не обладала никаким структурным сродством ни с субстратом, ни с продуктами реакции. Каким образом она может повлиять на каталитическую активность фермента. [2]
Молекула фермента состоит нз двух субъединиц ( мол. В состав активного центра входят также остатки гнстидина и по крайней мере 2 из 7 - 8 принадлежащих ферменту групп 5Н, к-рые необходимы для проявления каталнтич. Величина рН, при к-рой проявляется макс, каталитнч. [3]
Молекулы ферментов из-за их больших размеров ( большого молекулярного веса) невозможно или очень трудно вводить внутрь клеток. Для того чтобы повлиять на внутриклеточные биохимические процессы, необходимо применение низкомолекулярных веществ, которые могли бы легче проникать в клетки. Известны две группы таких веществ: а) предшественники активных групп ферментов - коферменты, витамины или их антагонисты ( антивитамины) и б) ингибиторы или активаторы ферментов. [4]
Молекулы ферментов, как и все белковые молекулы, построены из остатков а-аминокислот, соединенных пептидными связями. Линейную последовательность остатков в полипептидной цепи называют первичной структурой белка. Под вторичной структурой понимают характер спирализации или свертывания полипептидной цепи; эта структура стабилизируется водородными связями между карбонильной и амидной группами пептидных связей. В результате дальнейшего скручивания молекулы, уже имеющей определенную вторичную структуру, возникает третичная структура, которая стабилизируется за счет различных взаимодействий между боковыми группами аминокислот. Наконец, под четвертичной структурой понимают крупные белковые агрегаты, состоящие из нескольких полипептидных цепей различного типа. Помимо полипептидной цепи, на которую приходится основная масса молекулы, белок может содержать также и другие ковалентно связанные с полипептидной цепью химические группировки, называемые простетиче-скими группами. [5]
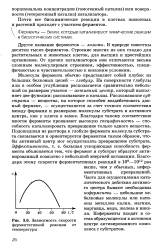 |
Зависимость скорости став образующегося в активном. [6] |
Молекула фермента обычно представляет собой клубок из больших белковых цепей - глобулу. На поверхности глобулы или в особом углублении располагается сравнительно небольшой по размерам участок - активный центр, который выполняет две функции: распознавание и катализ. Распознавание субстрата - вещества, на которое способен воздействовать данный фермент, - осуществляется за счет точного соответствия между формами и размерами молекулы субстрата и активного центра, как у ключа в замке. Благодаря такому соответствию многие ферменты проявляют высокую специфичность - способность катализировать превращение только одного вещества. Подошедшая из раствора к глобуле фермента молекула субстрата связывается и ориентируется ферментом таким образом, чтобы активный центр мог осуществлять превращение субстрата. [7]
Молекула фермента имеет различные химические группы, степень диссоциации которых изменяется в зависимости от кислотности или щелочности среды. Молекула фермента существует преимущественно в какой-то одной ионной форме в весьма узком интервале рН, и соответственно небольшая протяженность интервала, в котором большинство ферментов проявляет активность, указывает на то, что каталитической активностью обладает лишь одна из этих возможных форм. [8]
Если молекулы фермента и субстрата подходят друг к другу, то под влиянием электрических сил притяжения быстро формируется фермент-субстратный комплекс. [9]
Каждая молекула фермента в растворе несет некоторый суммарный заряд, от которого в известной сте пени зависит характер ее взаимодействий с другими макромолекулами; это взаимодействие не может быть точно предсказано на основании первичной структуры. [10]
Каждая молекула фермента содержит 1 атом магния. [11]
Участок молекулы фермента, вовлеченный в каталитический процесс. [12]
Способность молекулы фермента обеспечивать необходимое для данной реакции специфическое пространственное взаиморасположение реагирующих молекул обеспечивает резкое ускорение соответствующих реакций. Каталитическое действие ферментов обусловлено наличием в них активных участков. [13]
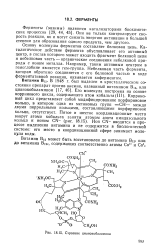 |
Строение цианокобаламина. [14] |
Основу молекулы ферментов составляет белковая цепь. Каталитическое действие фермента обусловливает его активный центр, в состав которого может входить фрагмент белковой цепи и небелковая часть - органические соединения небелковой природы с ионом металла. Например, таким активным центром в гемоглобине является гем-группа. Небелковая часть фермента, которая обратимо соединяется с его белковой частью в ходе ферментативной реакции, называется коферментом. [15]
Страницы: 1 2 3 4