Молекула - фермент
Cтраница 3
По одноцепочечному способу молекула фермента, атаковав в случайном порядке одну из полисахаридных цепей, последовательно отщепляет от нее звенья до тех пор, пока цепь полностью распадется. Лишь после этого фермент атакует следующие цепи. За время существования одного фермент-субстратного комплекса гидролизуются все доступные для фермента связи. [31]
Лишь после этого молекула фермента освобождается и атакует другую молекулу ( или фрагмент) субстрата. [32]
 |
Области применения ферментов в биологии и медицине ( по Грину. [33] |
Таким образом, молекула фермента характеризуется уникальностью структуры, которая и определяет уникальность ее функции. [34]
Именно так действуют молекулы дыхательных ферментов. У них, конечно, окисленная форма будет активно присоединять электроны, сразу попадающие на самый нижний уровень, а восстановленная легко отдавать электроны, находящиеся на самом верхнем энергетическом уровне. Именно поэтому такие молекулы хорошо выполняют функции переносчиков электронов. [35]
Точное знание конфигурации молекулы фермента, включая конформацию протеина в складках или при свертывании молекул в спираль, а также химической структуры вблизи активного центра может оказаться весьма важным. Так, например, активность фермента может быть нарушена при нагревании или воздействии химических реагентов ( кислот, оснований или концентрированных растворов солей), изменяющих конформацию протеина путем денатурации. Кроме того, активность может быть потеряна в результате действия ингибиторов. Последние обычно действуют при взаимодействии с активными центрами и используются для определения числа активных центров в отдельной молекуле фермента, а также для получения данных об их химическом строении. [36]
Согласно современным представлениям молекулы фермента и субстрата являются динамическими системами. Они меняют свою структуру в процессе создания комплекса фермент субстрат и в процессе его распада на фермент и продукты реакции. [37]
О пространственной структуре молекул ферментов известно пока мало. Считают, что количество спиральных участков в них невелико; это было доказано методом рентгеноструктурного анализа для ряда белков, в частности яичного альбумина; наиболее подробно - для лизоцима, а также для рибонуклеазы, а-химо-трипсина. Несомненно, что каждый ферментный белок обладает уникальной третичной структурой, где между спирализованными участками располагаются неспирализованные, количество которых велико и которые составляют значительную часть молекулы. Часто они удерживаются вместе с помощью атомов металлов, коферментов. Разбавление во многих случаях приводит к диссоциации субъединиц, иногда же силы, связывающие их, более прочны и для этого требуется действие концентрированных растворов мочевины. Химотрипсин, например, находится в растворах чаще в виде димера, при разбавлении образуется мономер ( Л1 - 23000), а в концентрированных растворах содержится тример. [38]
Общая особенность строения молекул ферментов заключается в сочетании разнообразия состочний, характерных для коферментных сопряженных систем с разнообразием положений, определяемым возможностями полипептидной цепи. Если вернуться к рассуждениям Эшби, о которых мы говорили в главе I, то нетрудно заметить, что ферментные аппараты в этом смысле наилучшим образом обеспечивают необходимое разнообразие состояний регулятора. Конструктор электронных реле и регулирующих устройств не может ограничиться только проводниками, его возможности, очевидно, будут шире, если он использует и изоляторы с надлежащей совокупностью геометрических и механических свойств. [39]
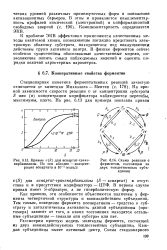 |
Кривые v ( S для аспартат-т-раис-карбамилазы. По оси абсцисс - концентрация аспартата в 10 - 3 моль / л.| Схема реакции с ферментом, состоящим из двух тождественных субъединиц. [40] |
Рассмотрим простую модель молекулы фермента, состоящей из двух взаимодействующих тождественных субъединиц. Каждая субъединица имеет активный центр. [41]
Рибофлавин входит в молекулу фермента в виде соединения с фосфорной, адениловой кислотами в виде кофермента. [42]
Здесь Е означает конформационно измененную молекулу фермента. [43]
Ди-сульфидные связи в молекуле фермента отсутствуют, так как протеи-наза содержит только один остаток полуцистина. [44]
Присоединение их к молекуле фермента происходит вне активного центра, но тоже приводит к уменьшению активности фермента. [45]
Страницы: 1 2 3 4