Бислой
Cтраница 1
 |
Температуры фазового перехода различных фосфолипидных бислоев. [1] |
Бислои, состоящие из одинаковых фосфолипидных частиц, при характеристической температуре претерпевают хорошо выраженный фазовый переход из относительно жесткого гелеобразного состояния в текучее жидкокристаллическое состояние. Помимо спектроскопических и дифракционных методов ( см. выше) для изучения такого фазового перехода применяют калориметрию. [2]
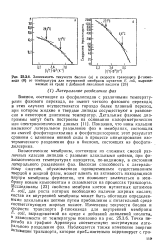 |
Зависимость текучести бислоя ( а и скорости транспорта [ 5-глюко-зида ( б от температуры для внутренней мембраны мутантов Е. coli, выращиваемых на среде с добавкой линолевой кислоты. [3] |
Бислои, состоящие из фосфолипидов с различными температурами фазового перехода, не имеют четкого фазового перехода в этих случаях осуществляется гораздо более плавный переход, при котором жидкие и твердые липиды сосуществуют в равновесии в некотором диапазоне температур. Показано, что ионы кальция вызывают латеральное разделение фаз в мембранах, состоящих из смесей фосфатидилхолина и фосфатидилсерина; этот результат сравним с влиянием ионной силы в процессе инициирования изотермического разделения фаз в бислоях, состоящих из фосфатидилсерина. [4]

Бислои ПАВ и амфифильных липидов являются ключевыми компонентами клеточных мембран. Данная чрезвычайная биологическая значимость делает понимание организации и ассоциации бислоев важнейшей и широко распространенной задачей. [6]

Схема бислоя клеточной мембраны ( жидкомозаичная модель): 1 - гидрофобные концы; г - гидрофильные головки липидов; 3 - молекулы холестерина; 4 - глобула белка; 5 - мопо - и олигосахариды. [8]
Гликолипиды образуют бислои, толщина и площадь которых ( в пересчете на молекулу) сходны с таковыми для фосфолипидов. Интересно отметить, что температура фазового перехода экстрагированных из мозга мясного скота цереброзидов составляет около 70 С из-за преобладания в них 24: 0 - и 24: 1-алкильных цепей; физиологическое значение такой высокой температуры фазового перехода не очень понятно. Температуры фазового перехода моно-и дигалактозилглицеринов из хлоропластов, напротив, лежат ниже С и, следовательно, при физиологической температуре эти липиды находятся в жидкокристаллической фазе. Разнообразие остатков, находящихся в области полярных головок гликолипидов, должно влиять на свойства клеточных поверхностей; например, групповая специфичность крови связана с гликопротеннами и гли-колиппдами мембраны эритроцитов. [9]
Самопроизвольное формирование бислоев было замечено в бинарных системах ПАВ-вода в оптически изотропной фазе, называемой губчатой. В данном случае, бислойные пленки ПАВ разделяют водную фазу на две части. Другая важнейшая область, в которой многие ПАВ формируют бислои, - это пены. Сток жидкости, разделяющей поверхности пузырьков ( воздух-вода), приводит к образованию пены и, как следствие, формированию бислоя. Так называемые черные пленки или черно-липидные мембраны формируются для изучения структуры и транспорта веществ посредством бислойных пленок. [10]
Уменьшение сопротивления бислоев при растворении в них молекул валиномицина, энниатина, монамицина и некоторых других исследовалось во многих лабораториях мира. Однако, как отмечается в работе [ 51, с. Антибиотики валиномицин, энниатин и монатицин представляют собой циклические депсипептиды, построенные из остатков аминокислот и оксикислот, соединенных между собой амидными и сложноэфирными связями. [11]
 |
Формирование кинков на углеводородной цеп и вследствие со пряженных гош / гранс-иэомериэацин.| Влияние фазового перекода на упаковку молекул в бислое. [12] |
В плоскости липидиого бислоя распределение липидных молекул может быть неоднородным в зависимости от состава липидоа, условий окружающей среды и действия различных мембранотроп-ных агентоа. Важный аспект гетерогенности липидов в бислое состоит в том, что перераспределение липидных молекул между различными доменами подразумевает их способность к легкой миграции вдоль поверхности бислоя или, иными словами, способность к так называемой латеральной диффузии. В жидкокристаллическом состоянии бислоя скорость латеральной диффузии липидных молекул очень высока. [13]
Динамическая структура лн-пидного бислоя, М, 1981, Бергельсон Л Д, Мембраны, молекулы, клетки, М, 1982, Ивков В Г, Берестовский Г Н, Липидный бислой биологических мембран, М, 1982, Кагава Я, Биомембраны, пер с япон, М, 1985, Сим Э, Биохимия мембран, пер с аигл, М, 1985, Болдырев А А, Введение в биохимию мембран, М, 1986, Биологические мембраны, под ред Дж Б С Финдлея и В X Эванза, пер с англ, М, 1990 Л И Барсуков. [14]
 |
Мозаичная модель клеточной мембраны. [15] |
Страницы: 1 2 3 4 5