Остатки - триптофан
Cтраница 3
Использование флуорометрических методов ограничивается тем, что далеко не все поглощающие ультрафиолетовое излучение вещества являются достаточно эффективными флуорофорами. Тем не менее среди них находятся такие аминокислоты, как триптофан и тирозин, в результате чего флуоресцируют все содержащие их белки. При облучении светом длиной волны 280 нм, т.е. в максимуме поглощения остатков триптофана, наблюдается флуоресценция с максимумом испускания при 348 нм. [31]
 |
Встраивание токсина В. thuringiensis в мембрану эпителиальной клетки кишечника насекомого и образование ионного канала. [32] |
Для биоконтроля численности насекомых-вредителей распыляют 1 5 109 - 2 5 109 спор В. Обработку проводят в тот период, когда число личинок в популяции насекомого-мишени максимально, поскольку параспоральные кристаллы чувствительны к солнечному свету и быстро разрушаются. В лабораторных условиях на свету за 24 ч разлагается более 60 % остатков триптофана в белках па-распорального кристалла, а в окружающей среде в зависимости от освещенности кристаллы могут сохраняться от одних суток до одного месяца. Такая нестабильность инсектицидного протоксина подразумевает, что появление устойчивых к нему насекомых маловероятно. [33]
Первичная структура грамицидина В, исследуемый образец которого содержал 89 % Уа - грамицидина В и 11 % Пеи грами-цидина В, выяснена Сарджесом и Виткопом [2765] путем 14-кратного ступенчатого расщепления дезформилграмицидина - В по методу Эдмана. Грамицидины В очень близки по своему аминокислотному составу к грамицидинам А; различие состоит лишь в замене остатка ь - Тгу12 - остатком L-фенилаланина. Грамицидин CD также построен аналогично грамицидину А, но со-держит остаток тирозина вместо одного из остатков триптофана. [34]
Для разделения небольших пептидов и аминокислот обычно используют смолы с 2 % дивинилбезола, реже с 4 и 8 % сшивающего агента. При попытках разделения на таких ионитах макромолекул, например белков или крупных пептидов, наблюдается сильная неспецифическая адсорбция за счет вандерваальсовых сил или электростатического взаимодействия заряженных группировок на поверхности глобулы с ионообменными центрами матрицы. Для небольших молекул ситовой эффект не играет существенной роли. Некоторые пептиды, особенно содержащие остатки триптофана, необратимо сорбируются полимерной матрицей. [35]
Фермент представлен двумя изоформами с мол. Фермент имеет 2 активных центра с одинаковой каталитич. В состав активного центра входят остатки триптофана, гистидина, аргинина и карбоксильная группа. [36]
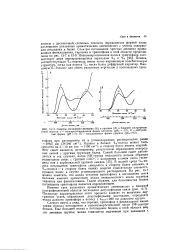 |
И. Спектры поглощеии цитидина ( А и уридина ( Б в товой области, / - монопротонированная форма цитидина ( р / ( а 4 2. [37] |
Однако в белках La-полоса может быть смещена на 3 - 10 нм ( на - 1100 см-1) в сторону более низких энергии. Этот сдвиг является, по-видимому, результатом образования водородных связей с другими группами белка. В водной среде U - no - лоса поглощения триптофана смещается в сторону более высоких, а La-полоса-в сторону более низких энергий относительно соответствующих полос в случае углеводородного растворителя. Из рис. ю / можно заметить, что вклад остатков триптофана в поглощение света белками намного превосходит вклад эквивалентного числа остатков тирозина или фенилаланина. Таким образом, для большинства белков поглощение триптофана является определяющим. [38]
При использовании этого реагента дальнейшее окисление вообще не идет даже при высоких концентрациях Н2О2, поэтому таким путем определяют четкие изобестические точки. Применяющийся для этой цели диоксан должен быть очищен от более реакционноспособных органических перекисей. В некоторых белках остатки триптофана окисляются этим реагентом ступенчато. По-видимому, это указывает на скачкообразные конформационные изменения белковой молекулы в процессе окисления остатков триптофана. [39]
Как известно, функция рибонуклеазы состоит в гидролитическом расщеплении рибонуклеиновых кислот и олигонуклеотидов. Как мы видели, это один из первых белков, изучавшихся с помощью ЯМР, хотя спектры, полученные на ранних стадиях, не обнаруживали характерных деталей. Рибонуклеаза близка по размеру ( молекулярная масса 13700, 124 аминокислотных остатка) и форме к лизоциму и является удобным объектом для изучения методом ЯМР. Таким образом, в растворе при нейтральных рН молекула заряжена положительно. В дополнение к 4 Гис имеются также 6 Тир и 3 Фен, но нет остатков триптофана. Форма ее глобулы близка к сферической; имеется большая щель, в которой происходит связывание субстрата. С одной стороны этой щели расположены в непосредственной близости друг от друга остатки Гис-12, Гис-119 и Лиз-7, а с другой стороны находится Лиз-41. По данным подробных химических исследований все эти четыре остатка входят в активный центр. [40]
Два графика, полученные таким образом, выявляют тонкую структуру спектра. Преимуществом указанного метода служит то, что проведенная с помощью ЭВМ аппроксимация кривой дает представление о форме самой полосы поглощения. Как нетрудно видеть, связывание сукцината и карбамоилфосфата вызывает слабый сдвиг ( 20 см - [) полосы поглощения и очень небольшое ее уши-рение. Основной эффект состоит в более четком выражении колебательной структуры 0 - 0-полосы при 34 600 см -, обусловленной поглощением двух остатков триптофана, которые присутствуют в субъединице фермента. Причина этого изменения, однако, не вполне понятна, в чем и проявляется ограниченность метода разностной спектроскопии. [41]
В яде Hydrophiidae ферментов мало, зато в нем много нейротоксинов, обладающих курареподобным действием. Эти нейротоксины представляют собой полипептиды основного характера, в состав которых входит от 60 до 74 аминокислот. Сходные нейротоксины содержатся и в ядах Е1а - pidae. Для биологической активности этого токсина необходимо наличие четырех ди-сульфидных мостиков между остатками цистеина. По-видимому, щелочные аминокислоты токсина необходимы для его избирательного связывания с рецептором. Остатки триптофана и тирозина также играют важную роль, удерживая молекулу кобротоксина в конформации, соответствующей максимуму биологической активности. [42]
Все эти остатки способны к вандерваальсовым взаимодействиям и обладают сильно выраженными гидрофобными свойствами. Фенольная группировка остатка тирозина способна, помимо того, участвовать в качестве донора в образовании водородной связи. Индольное кольцо триптофана обладает рядом интересных и совершенно особых свойств. Это единственный остаток в белковой молекуле, способный очень легко вступать в реакцию образования комплекса с переносом заряда в качестве донора. Соединения индола являются очень хорошими донорами электронов при образовании комплексов с переносом заряда с соединениями пиридиния и другими электрофильными соединениями. Остатки триптофана определяют практически всю ультрафиолетовую флуоресценцию белков. Это верно даже в отношении белков, содержащих много тирозина и мало триптофана, хотя остатки тирозина ( и в меньшей степени фенилала-нина) дают вклад в ультрафиолетовое поглощение белков. Как поглощение, так и флуоресценция остатков триптофана в ультрафиолетовой области весьма чувствительны к изменениям их ближайшего окружения в молекуле белка. Пиррольный азот триптофана может служить донором в некоторых реакциях образования водородной связи. [43]
С) осадок окрашивается сначала в розовый или желтый, а затем в пурпурно-красный цвет. Красное окрашивание обусловлено образованием ртутных солей нитрофенолов, благодаря присутствию в белке остатков тирозина. Присутствие в белке остатков триптофана дает буро-красное окрашивание. [44]
Между синглетными состояниями хромофоров перенос электронной энергии возбуждения возможен на расстояния порядка 30 А, тогда как триплет-триплетный перенос энергии требует значительно меньших расстояний - около 12 А. Если синглетное состояние триплетного акцептора обладает большей энергией, чем синглетное состояние триплетного донора, последний может возбуждаться светом с длиной волны, не поглощаемым триплетным акцептором. В этих условиях фосфоресценция триплетного акцептора наблюдается только при триплет-триплетном переносе энергии. Фосфоресценция триптофана карбоангидразы вызывается светом с Л ] 280 нм. Фосфоресценция сульфамида возбуждается светом с длиной волны 330 нм, которая яе вызывает свечения фермента. Следовательно, остатки триптофана расположены очень близко от центра связывания, образуя, видимо, поверхность его полости. [45]
Страницы: 1 2 3 4