С-концевые остатки
Cтраница 1
С-Концевые остатки аргинина можно заменять на остатки других основных или структурно близких аргинину аминокислот; в этом случае наблюдается лишь небольшое снижение активности. Аналогичная замена на нейтральные аминокислоты сопровождается резким снижением активности. Значительным снижением активности сопровождается одновременная замена двух остатков аргинина. К такому же результату приводит замена остатков фенил-аланина ( положения 5 и 8) на остатки неароматических аминокислот. Замена в положении 6 ( остаток серина) практически может происходить без изменения активности. [1]
С-концевых остатков при В - и R-формах не превышают 1 0 ккал / моль. [2]
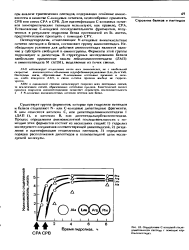 |
Определение С-концевой последовательности пептида с помощью кар-боксипептндаз. [3] |
С-концевых остатков, целесообразно применять СРВ или смесь СРА и СРВ. Для идентификации С-концевых остатков химотриптических пептидов используют, как правило, СРА. [4]
С-концевых остатков, состав ( Gly-X-V) m в ней строго соблюдается. [5]
Определение С-концевых остатков в пептидах и белках разработано менее удовлетворительно, нежели определение Af-концевых. [6]
Для определения С-концевых остатков чаще всего используют ферментативный гидролиз карбокснпептидазами, к-рые специфически расщепляют пептидные связи, образованные С-коицевыми остатками. Поскольку после отщепления концевых остатков фермент атакует послед, пептидные связи, измерение скорости отщепления отдельных аминокислот позволяет анализировать также и С-концевую аминокислотную последовательность. [7]
Карбоксипептидаза последовательно отщепляет от пептидов С-концевые остатки. В результате последовательного действия этих протеолитических ферментов и пептидаз перевариваемые белки в конечном итоге превращаются в смесь свободных аминокислот, которые далее транспортируются через эпителиальные клетки, выстилающие тонкие кишки. Свободные аминокислоты проникают в капилляры ворсинок и переносятся кровью в печень. [8]
Действительно, ферментативное отделение четырех С-концевых остатков лишает стабильности четвертичную структуру дезоксигемоглобина и ингибирует гем-гем взаимодействие. Энергия одного солевого мостика равна 1 - 2 ккал / моль. [9]
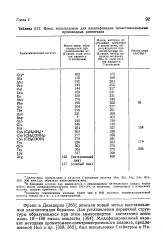 |
Ионы, используемые для идентификации триметилсилильньпс производных дипептидов. [10] |
Дипептиды, содержащие в качестве С-концевых остатки Abu, Ala, Arg, Gin, Glu, Oly, Met или Lys, образуют дикетопиперазины. [11]
Известные в настоящее время методы определения С-концевых остатков менее удовлетворительны. Карбоксипептидаза действует только на свободные а-карбоксильные группы, подобно тому как лейцинаминопептидаза действует на свободные а-аминогруппы. Другой метод, разработанный Акабори, основан на полном гидразинолизе полипептида. Гидразин реагирует с каждой пептидной связью, превращая все аминокислоты ( за исключением С-кон-цевой, карбоксил которой не участвует в образовании пептидной связи) в ацилгидразины. С-концевую аминокислоту отделяют и идентифицируют хроматографически. Значительно реже используется метод, основанный на восстановлении С-концевой карбоксильной группы до спиртовой с помощью подходящих доноров гидрид-иона. При последующем полном гидролизе полипептида образуется аминоспирт, соответствующий С-концевой аминокислоте. Этот аминоспирт может быть выделен и идентифицирован. [12]
Пептидные связи расщепляются с образованием гидразидов аминокислот, а С-концевые остатки отщепляются в виде свободных аминокислот. [13]
Все три перечисленных метода полностью могут быть использованы и для идентификации С-концевых остатков. Однако относительное их значение для практических применений весьма различно в обоих случаях. [14]
ХТГ - химотрипоиноген А; Нео-ХТГ - неохимотрипсиноген; 6 - ХТ и - ХТ - соответствснБ: о S-и а-химотрипсин; РТ - реакции, катализируемые трипсином; РХТ - реакции, катализируемые химотрипоином; и - соответственно N - и С-концевые остатки; А, В и С - А -, В - и С-цепи а-химо-трипсина; Асп - остаток аспарагина. Две цепи в молекулах неохимотрипсиногена и 6-химотрипсина и три цепи в а-химотрипсине связаны между собой дисупьфидными мостиками. [15]
Страницы: 1 2 3