С-концевые остатки
Cтраница 2
В некоторых стучвях тритий включается в остатки аспарагиновой и глутами-новой кислот, расположенные в середине пептидной цепи. С-Концевые остатки пролииа в описанных условиях не образуют оксазолона, в в С-кониевые остатки треонина и сернна обычно не удается ввести достаточного количества радиоактивной метки, что, вероятно, связано с деградацией последних в присутствии уксусного ангидрида. [16]
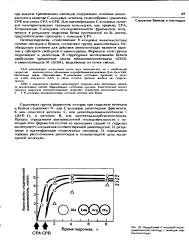 |
Определение С-концевой последовательности пептида с помощью кар-боксипептндаз. [17] |
С-концевых остатков, целесообразно применять СРВ или смесь СРА и СРВ. Для идентификации С-концевых остатков химотриптических пептидов используют, как правило, СРА. [18]
Поскольку изменение положения С-концевых остатков приводит к разрыву солевых мостиков с другими субъединицами, необходимое изменение третичной структуры одной субъединицы переходит в небольшое изменение четвертичной структуры всего белка. Далее небольшие изменения четвертичной структуры, как и другие изменения в субъединицах, окончательно индуцируют большой структурный переход, состоящий в изменении четвертичной структуры от R - к Т - форме. Другими словами, процессы, в которых участвует железо в каждой отдельной субъединице. [19]
Нужно отметить, что специфичность карбоксипептидазы по отношению к субстрату в значительной мере определяется природой С-концевого остатка. Большинство эффективно расщепляемых субстратов имеет ароматические С-концевые остатки, тогда как концевой пролин блокирует пептид от действия карбоксипептидазы. У а-кортикотропина освобождаются только три аминокислоты, потому что четвертой аминокислотой является пролин. [20]
Тройные спирали коллагена большинства позвоночных состоят из двух о - цепей и гомологичной 2-цепи. За исключением 16 N-концевых и 25 С-концевых остатков, состав ( Gly-X-V) m в ней строго соблюдается. Пользуясь этой формулой, можно легко выявить в различных аминокислотных последовательностях коллагеноподобные структуры. В глобулярных белках такие структуры пока еще не быт обнаружены. Однако весьма вероятно, что они присутствуют в компоненте Clq системы комплемента человека [ 1981, которая узнает антитела, соединенные с антигенами. Как установлено по электронным микро-фотограммам, это белок содержит пучок из 18 параллельных цепей, организованных в шесть коллагеноподобных волокон, которые входят в шесть глобул. Возможно, что в дальнейшем будут выявлены и другие смешанные белки, содержащие коллагеноподобные структуры. [21]
Этим реакциям явно благоприятствует невысокая скорость образования нужной пептидной связи, что происходит из-за пространственных препятствий, и по этой причине объемистые аминокислотные остатки, например валин и изолейцин, не следует располагать в местах сшивки. Во всех случаях, когда пептидные фрагменты содержат С-концевые остатки, не являющиеся глицином или пролином, должна использоваться также методика, делающая риск рацемизации минимальным. [22]
Этому препятствует, однако, тот факт, что С-концевые остатки мио-глобина образуют часть спирали Н и потому не могут так легко менять свое окружение. [23]
Пауэре ( личное сообщение) установил, что гемоглобины без эффекта Бора отличаются от обычных концевыми аминокислотами ос - и Р - цепей. Известно, что у млекопитающих за эффект Бора ответственны главным образом С-концевые остатки гистидина в р-цепях и свободные N-концевые аминогруппы а-це-пей. В одном гемоглобине рыб, не обнаруживающем эффекта Бора, С-концевой аминокислотой Р - цепей оказался не гистидин и, кроме того, N-концевые группы - цепей были блокированы. [24]
Специфичность обеих экзопептидаз различна. Карбоксипептидазой А, впервые использованной Ленсом в 1949 г. для определения С-концевых остатков в инсулине, отщепляются все аминокислоты, кроме Lys, Arg, Pro-и His. Карбоксипептидаза В отщепляет Lys, Arg, Orn и S-аминоэтилцистеин. Оба фермента взаимно дополняют друг друга и, как правило, применяются в комбинации. [25]
Реакции конденсации, протекающие с участием карбоксильной труппы, не нашли широкого применения [393], но реакции, при которых происходят превращения этой группы, были подробно исследованы. Недавно было установлено, что фенилмагаий-бромид превращает эфирную - пруту эфира пептида в кетой [393] и что в ходе реакции разложения Бергмана - Зерваса [ 119а ] ( стр. С-концевые остатки превращаются в соответствующие низшие альдегиды. Однако эти реакции представляют только академический интерес. [26]
Наряду с этим, если те же самые остатки располагаются внутри пептидной цепи, они не изменяются в ходе восстановления и обнаруживаются после гидролиза в виде аспарагиновой и глутаминовой кислот. Остатки же аспарагиновой и глутаминовой кислот в пептидной цепи будут подвергаться восстановлению, если их со-карбоксильные группы этерифпцированы; при этом будет образовываться у-а - ОАМ и соответственно б-а - ОАВ. Наконец, С-концевые остатки аспарагн-новой и глутаминовой кислот при восстановлении будут давать диолы. Чнбнол и др. [29] разработали метод определения амидных групп в белках, основанный на использовании боргидрида лития и применении периодатного окисления 1 2-оксиаминов и диолов. [27]
При кислотном гидролизе обработанного таким образом материала остатки аспарагино-вой кислоты дают лактон гомосерина, а остатки глутаминовой кислоты появляются в виде а-амино-8 - оксивалерьяновой кислоты. Остатки глутамина и аспарагина, присутствующие в молекуле исходного белка, при гидролизе превращаются в соответствующие дикарбоновые кислоты. Один из способов определения С-концевых остатков основан на применении карбокси-пептидазы ( из сока поджелудочной железы), специфически отщепляющей С-концевую аминокислоту. Этот метод страдает рядом недостатков, один из которых состоит в том, что фермент может отщеплять наряду с С-концевой группой следующий за ней аминокислотный остаток. [28]
Присоединение субстрата превращает заполненную молекулами воды полость в сравнительно гидрофобный участок. Остаток Leu-203 выступает внутрь полости, тогда как в свободном ферменте он взаимодействует с другими аминокислотами, образующими р-структуру. Однако вода удаляется из полости лишь частично, и три или четыре ее молекулы остаются внутри после присоединения тирозильной группы субстрата. Поскольку в этом участке происходит связывание боковых групп гидрофобных С-концевых остатков и субстраты с такими остатками гидролизуются быстрее всего ( рис. 15.2), то эту полость можно назвать участком специфичности. При этом надо иметь в виду, что другие участки связывания также влияют на специфичность. [29]
Положительно заряженные остатки аргинина взаимодействуют с С-концевой карбоксильной группой. Это взаимодействие является главным фактором, обеспечивающим селективность карбоксипептидазы именно к С-концевым аминокислотам. Гидрофобные компоненты активного центра обеспечивают преимущественную фиксацию в нем гидрофобных С-концевых остатков. Уже из этого рассмотрения становится ясной абсолютная стереоспецифичность фермента - при аналогичной посадке D-амино-кислотного остатка в сторону каталитического центра - координированного иона Zn - будет направлен атом Н при хиральном атоме С. Дополнительно стереоспецифичность обеспечивается формой полости, в которой происходит фиксация субстрата. Внутренняя поверхность полости в направлении связанного с хираль-ным центром С-концевой аминокислоты атома Н достаточно близка к нему и не допускает замены его на какой-либо превышающий его по размеру фрагмент без существенной деформации полости. Поэтому даже связывание С-концевого фрагмента в D-конфигурации затруднено. [30]
Страницы: 1 2 3