Остаток - гистидин
Cтраница 4
При пептидных синтезах с незащищенной имидазольной функцией могут встречаться трудности, связанные с возможностью ацилирования имида-зольиого остатка, его слабой основностью, а также часто с низкой растворимостью таких пептидов. Некоторые синтезы с незамещенной имидазольной функцией тем не менее были описаны, причем остаток гистидина был аминокомпонентом. [46]
Таким образом, происходящая реакция сводится к образованию промежуточного ацилфермента, а затем к переносу ациль-ного остатка к молекуле воды, если протекает гидролиз, или к иному акцептору ацетильной группы. Серии, конечно, содержится в активном центре не всех ферментов. Так, например, у ри-бонуклеазы его нет, и в этом ферменте, как предполагается, два остатка гистидина выполняют функцию акцептора и донора протонов. Серии и гистидин, несомненно, являются не единственными остатками, необходимыми для работы активного центра. Имеются факты, говорящие об участии триптофана в активных центрах химотрипсина, трипсина, лизоцима, однако об этом пока еще известно мало. [47]
Дэви и др. [4] после действия фермента при рН 7 6 удалось выделить в кристаллической форме производное ( З - лактоглобулина, у которого были удалены оба С-концевых остатка изолейцина. Далее воздействием карбоксипептидазы при рН 9 2 получили другое кристаллическое производное, у которого были удалены оба концевых остатка гистидина. [48]
ИЗОЦИТРАТ-ЛИАЗА ( / лрео - О-изоцитратглиоксилат-лиа - за), фермент класса лиаз, катализирующий обратимое расщепление изолимоииой ( 1-гидрокси - 1 2 3-пропантрикар-боновой) к-ты до глиоксиловой и янтарной. Играет важную роль в глиоксилатном цикле. На С-концах полипептидных цепей локализован остаток гистидина, на М - коицах-метионина. Каждая субъединица содержит 21 остаток цистеина, 5 из к-рых важны для ферментативной активности. [49]
Значительные усилия были потрачены на установление природы лигандов атома цинка до того, как стали известны трехмерная структура и аминокислотная последовательность КПА. В результате был сделан вывод, что атом цинка связан с цистеином и а-аминогруппой. На самом деле лигандами цинка являются остаток глутамата и два остатка гистидина. Было бы полезно для будущего проанализировать некоторые опасности, подстерегающие при химическом определении природы лигандов. [50]
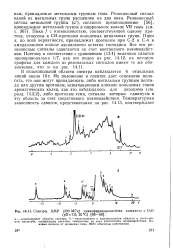 |
Спектры ПМР ( 220 МГц цианоферримиоглобина кашалота в D2O. [51] |
Резонансный сигнал одной из метальных групп расщеплен на два пика. Резонансный сигнал метальной группы ( а), согласно предположению [56], принадлежит метильной группе в пиррольном кольце VR тема ( см. с. Пики / с интенсивностью, соответствующей одному протону, отнесены к СН-протонам кольцевых винильных групп. Пики х, по всей вероятности, принадлежат протонам при С-2 и С-4 в имидазольном кольце аксиального остатка гистидина. Все эти резонансные сигналы сдвигаются за счет контактного взаимодействия. [52]
Обратимая реакция гидратации - дегидратации фумарат малат катализируется ферментом фумаразой. Были тщательно исследованы кинетика и механизм действия кристаллической фумаразы, выделенной из сердца свиньи. Этот фермент с молекулярным весом 2 2 - 105 состоит из четырех одинаковых полипептидных субъединиц ( мол. Их значения рК при ионной силе 0 01 равны 6 2 и 6 8 соответственно; по-видимому, это либо два гистидино-вых остатка, либо один остаток гистидина и один остаток аминодикарбоновой кислоты. Реакция стереоспецифична в обоих направлениях. Добавление или отщепление элементов молекулы воды осуществляется только в транс-конформации. [53]
Структурные данные четко свидетельствуют о том, что стереохимические изменения железо-порфирина в гемопротеинах связаны с переходами спинового состояния центрального координированного иона металла. Однако из-за отсутствия достаточно точных стереохимических данных невозможно сделать каких-либо количественных оценок относительного вклада изменений структуры железопорфирина по сравнению с постулированным изменением длины связи Fe-Мимидазол. Несомненно, таким количественным оценкам препятствует тот факт, что дезоксигемоглобин и координированные тетрамерные гемогло-бины млекопитающих кристаллизуются в различных пространственных группах симметрии [113], вследствие чего невозможно прямое сопоставление соответствующих четвертичных структур, например, разностным методом Фурье. Кроме того, было постулировано [103, 128], что оба конформационных события носят три-герный характер и инициируют изменение четвертичной структуры при отщеплении или связывании молекулы кислорода вследствие изменений в F-спирали субъединиц белка, содержащих координирующий остаток гистидина. Предполагается, что это изменение структуры распространяется, на другие области белка. Однако природа изменений в F-спирали пока не ясна. [54]
Страницы: 1 2 3 4