Аминокислотный остаток
Cтраница 1
Аминокислотный остаток со свободной аминогруппой ( в природных белках это - а-аминогруппа) называется N-концевым; остаток, содержащий свободную карбоксильную группу, носит название С-концевого. Название полипептида составляется из названий входящих в его состав аминокислот, которые перечисляются последовательно, начиная с N-концевого остатка. [1]
Аминокислотный остаток со свободной аминогруппой ( в природных белках это - а-аминогруппа) называется N-концевым; остаток, содержащий свободную карбоксильную группу, носит название С-концевого. Название полипептида составляется из названий входящих в его состав аминокислот, которые перечисляются последовательно, начиная с N-концевого остатка. При этом суффикс ин в названиях аминокислот заменяется суффиксом ил, и только С-концевая аминокислота сохраняет суффикс ин. [2]
Аминокислотный остаток, химическая группировка строения - HN-CHR-СО -, являющаяся мономерным звеном белковой цепи. То, что остается от аминокислоты, когда она встраивается в белковую цепь. [3]
Аминокислотный остаток П, несущий своб, й-аминогруппу, наз. [4]
Аминокислотный остаток Lys-296, ответственный за связывание ретиналя, расположен ближе к С-концу белка. [5]
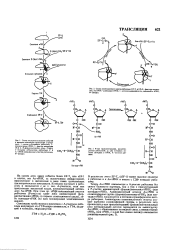
Аминокислотный остаток Аа - тРНК и сложноэфирная группа ( формил) метионил-т РНКр ( или пеп-тидил - тРНК) локализуется в пептидил-трансферазном центре рибосомы. [7]
Далее аминокислотный остаток переходит в молекулу специфической для данной аминокислоты растворимой т - РНК, причем аминокислота аци-лирует один из двух вторичных гидрокислов рибозы концевого нуклео-зида т - РНК. Это видно из того, что транспортная РНК, связавшая молекулу аминокислоты, в отличие от свободной т - РНК не окисляется йодной кислотой, как все z / ие-гликоли ( кн. I, стр. [8]
Присоединение каждого аминокислотного остатка к растущей полипептидной цепи происходит в три стадии. Этот цикл повторяется столько раз, сколько остатков следует присоединить. [9]
При замене аминокислотного остатка в пептиде название аминокислоты, которая замещает другую, а также положение замены указывают в квадратных скобках перед тривиальным названием соответствующего пептида. В краткой форме, которую применяют только в таблицах, место замены указывается цифрой, стоящей сверху у символа заменяющей аминокислоты. [10]
Кодон второго аминокислотного остатка должен начинаться на U, т.е. это могут быть Phe, Leu, Ser, Туг, Cys, Trp. И лишь третий аминокислотный остаток может быть любым. [11]
Успешное введение аминокислотного остатка гистидина в синтетические пептиды по-прежнему представляет собой чрезвычайно сложную проблему. И это связано с крайне неудобными для синтеза химическими свойствами имидазольного цикла. Свободный имидазол - это эффективный катализатор гидролиза сложных эфи-ров и амидов, а также рацемизации. Сами же гистидиновые производные особенно склонны к рацемизации в процессе пептидного синтеза. Если имидазольный цикл оставить незащищенным, то он может подвергаться ацилированию активированными карбоксильными компонентами, причем получающиеся ацильные производные сами по себе достаточно реакционноспособны и могут затем вызывать перенос ацильной группировки в разных участках молекулы. По этой причине Л т-ацильные производные гистидина часто неудобны в качестве синтетических интермедиатов, если на ряде стадий нужно сохранить находящуюся в боковом радикале защитную группу. Для ступенчатого синтеза можно использовать защищенные уретановые производные, например Na, N - m бис-грег-бут-оксикарбонилпроизводное ( 63), причем обе защитные группы удаляют непосредственно после введения аминокислотного остатка в пептидную цепь. [12]
Более того, акцепторный аминокислотный остаток атакует донорный остаток в такой ориентации, что реакция транспептидации дает в результате удлиненный остов N, - С. [13]
Крика, структура аминокислотного остатка не имеет отношения к выбору аминоацил-т РНКЧкодоном. Кодон комплементарно взаимодействует лишь с самой тРНК как адаптером, и, следовательно, тот аминокислотный остаток, который был присоединен к адапторной тРНК заранее, оказывается автЬмати - чески навязанным рибосоме, никак не участвуя в узнавании кодона. [14]
Действительно, если каждый аминокислотный остаток ( пептидная единица) может иметь три конформации R, В и L, то в декапепти-де возможно уже З10 конформации. В циклических пептидах благодаря условиям замыкания число возможностей несколько меньше; гораздо меньше оно для симметричных циклических пептидов, и потому в ряде случаев поиск оптимальной структуры становится технически выполнимым. [15]
Страницы: 1 2 3 4