Фосфатный остаток
Cтраница 2
При отщеплении от 1 7-дифосфата седогептулозы одного фосфатного остатка и остатка гликолевого альдегида получается 5-фосфат рибозы, изомери-зующийся в 5-фосфат рибулозы. Фосфорилирование последнего дает 1 5-ди-фосфат рибулозы. [16]
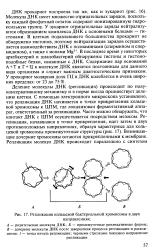 |
Репликация кольцевой бактериальной хромосомы в двух. [17] |
Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидро-ксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками - ги-стонами. В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоно-подобные белки, связанные с ДНК. Содержание пар оснований А ТиГ Цв молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. [18]
В течение ряда лет большинство исследователей считало, что фосфатный остаток расположен у аксиальной ОН-группы при С2 миоинозита, так как при гидролизе монофосфоинозитида был выделен 2-фосфат миоинозита. [19]
В отличие от этих макроэргических связей энергия связи третьего фосфатного остатка невелика. [20]
Энергия для транспортирования веществ может высвобождаться за счет отщепления фосфатного остатка от АТФ и, при действии дегидрогеназ. [21]
В отличие от этих макроэргических связей энергия связи третьего фосфатного остатка невелика. [22]
Одно включает наименование нуклеозида с указанием положения в нем фосфатного остатка ( например, аденозин - З - фосфат, ури-дин-5 - фосфат), другое строится с добавлением суффикса - овая кислота к названию остатка пиримидинового ( например, 5 -уриди-ловая кислота) или пуринового ( например, З - адениловая кислота) оснований. [23]
Сначала при элиминировании пирофосфата образуется смешанный ангидрид аминокислоты и фосфатного остатка аде-ниловой кислоты, который существует только в виде комплекса с ферментом. [24]
Действие рибонуклеазы направлено в основном на разрыв связи, соединяющей фосфатный остаток при С-3 в пиримидиннуклео-тиде с С-5 следующего нуклеотида. Поэтому рибонуклеазу можно рассматривать как высокоспецифическую фосфодиэстеразу, гидро-лизующую только вторичные фосфорные группы пиримидин-нуклеозид - З - фосфатов. В соответствии с этим фермент может также гидролизовать циклические 2 3 -вторичные фосфаты пиримиди-нуклеозидов. [25]
Кроме указанного выше ограничения - невозможности использовать карбодиимид для активации фосфатного остатка, находящегося рядом с незащищенной гидроксильной группой, имеется второе ограничение, не позволяющее использовать этот реагент для получения любых нуклеотидо - ( P-N) - пептидов. Следовательно, карбодиимидный метод не может быть использован для синтеза соединений типа III - аминокислотных производных по меж-нуклеотидному фосфору. Этого очень существенного ограничения лишен другой распространенный метод активации фосфатных остатков в ну-клеотидах - метод смешанных ангидридов, частным случаем которого ЯЕЗляется пирофосфатный метод. [26]
При осуществлении синтеза этих соединений следует считаться с возможностью миграции фосфатного остатка [189], а также с ацильной миграцией [190] при использовании сложноэфирных защитных групп в миоинозитном кольце. К защитным группам предъявляется ряд требований: возможность селективного введения, удаление без разрыва фосфоэфирных связей и с сохранением функциональных групп в глицериновой половине молекулы. [27]
Ввиду того что в положениях 2 и 3 легко происходит миграция фосфатного остатка, вопрос об избирательном фосфорилировании гидроксильнои группы в одном из этих положений остается открытым. [28]
В воде фосфорная кислота диссоциирует на три положительно заряженных иона водорода и один трехзарядный отрицательный фосфатный остаток. [29]
На основании данных кислотного и щелочного гидролиза можно сделать вывод, что в природных фосфоинозитидах фосфатный остаток присоединен к Ci молекулы миоинозита. Аналогичные выводы получены в результате превращения миоинозит-1 - фосфоглицерина в миоино-зит-1 - фосфат в условиях, почти исключающих фосфатную миграцию. [30]
Страницы: 1 2 3 4