Полиплоидизация
Cтраница 2
Подводя итоги современным данным по кариологии покрытосеменных растений, можно констатировать следующую закономерность: где преобладают мелкие хромосомы, там в процессе видообразования большую роль играла полиплоидия и межвидовая, а вероятно, и более отдаленная гибридизация с последующей полиплоидизацией геномов; путь видообразования через дислокацию генетического материала ( разнообразные хромосомные перестройки) характерен для видов с крупными хромосомами, тогда как полиплоидизация геномов здесь отсутствует или выражена слабо. Необходимо, однако, отметить, что не всегда наличие мелких хромосом обусловливает возможность появления полиплоидов. Известный, экономически важный, род Ribes - смородина характеризуется очень мелкими хромосомами и тем не менее полиплоидных видов в нем не обнаружено и, более того, экспериментальная полиплоидия не имеет успеха. [16]
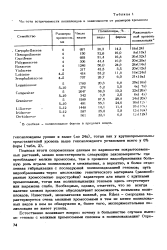 |
Частота встречаемости полиплоидов в зависимости от размеров хромосом. [17] |
Подводя итоги современным данным по кариологии покрытосеменных растений, можно констатировать следующую закономерность: где преобладают мелкие хромосомы, там в процессе видообразования большую роль играла полиплоидия и межвидовая, а вероятно, и более отдаленная гибридизация с последующей полиплоидизацией геномов; путь видообразования через дислокацию генетического материала ( разнообразные хромосомные перестройки) характерен для видов с крупными хромосомами, тогда как полиплоидизация геномов здесь отсутствует или выражена слабо. Необходимо, однако, отметить, что не всегда наличие мелких хромосом обусловливает возможность появления полиплоидов. Известный, экономически важный, род Ribes - смородина характеризуется очень мелкими хромосомами и тем не менее полиплоидных видов в нем не обнаружено и, более того, экспериментальная полиплоидия не имеет успеха. [18]
Полиплоидизация может быть двух типов: митртическая к мейотическая. Митотическая полиплоидизация происходит в соматической ткани и сразу приводит к возникновению клетки с удвоенным набором хромосом. При мейотической полиплоидиза-ции имеет место блока да движения хромосом в первом или во-втором делении мейоза, что приводит к образованию половых клеток с нередуцированным числом хромосом. Соединение двух таких гамет дает зиготу с удвоенным числом хромосом, из которой развивается тетраплоидный организм. Однако гораздо чаще нередуцированные гаметы сливаются в процессе оплодотворения с нормальными гаплоидными, давая начало триплоид-ному организму. [19]
Положительные результаты дает обработка органов взрослых растений методом инъекций. Для пшеницы разработан метод полиплоидизации путем инъекции 0 1 - 1 0 % - ного раствора колхицина в цветки на трех стадиях развития: до опыления, во время опыления и после опыления. Тетраплоидные формы у винограда получены при введении раствора колхицина в побеги на различных стадиях развития, начиная от только что раскрывшихся почек до молодых побегов длиной 5 см. Инъекция раствора колхицина в почки маточных растений капусты на самых ранних стадиях их развития оказалась очень эффективным приемом в индуцировании полиплоидии у этой культуры, позволившим значительно повысить образование полностью тет-раплоидных, не химерных растений. Для инъекции в этом случае применяют 0 25 % - ный раствор колхицина. [20]
Для успешного индуцирования полиплоидии далеко не безразлично, какую часть растения и в какой фазе развития подвергать обработке. В этом отношении большое преимущество представляет полиплоидизация формирующихся гамет, при последующем объединении которых возникают чисто тетраплоидные организмы, или полиплоидизация зиготы во время первого деления, полностью устраняющая трудности, связанные с выращиванием растений, состоящих из клеток с удвоенным и неудвоенным числом хромосом. [21]
Для успешного индуцирования полиплоидии далеко не безразлично, какую часть растения и в какой фазе развития подвергать обработке. В этом отношении большое преимущество представляет полиплоидизация формирующихся гамет, при последующем объединении которых возникают чисто тетраплоидные организмы, или полиплоидизация зиготы во время первого деления, полностью устраняющая трудности, связанные с выращиванием растений, состоящих из клеток с удвоенным и неудвоенным числом хромосом. [22]
При обсуждении роли полиплоидии в эволюции растений мы должны учитывать следующие обстоятельства. Полиплоиаизация хромосомного набора вида приводит не только к увеличению дозы соответствующего гена, что само по себе дает значительный фенотипический эффект и может иметь непосредственное адаптивное значение, но при дальнейших изменениях генов в результате мутагенеза полиплоидизация приводит к созданию новой генетической системы. Таким путем могут развиваться, вероятно, комплементарные и полигенные системы, имеющие определенное адаптивное значение. Известно, что каждый генокон - тролируемый процесс осуществляется системой, состоящей из многих генов. Продолжительность периода, в течение которого происходят метаболические процессы, также зависит от числа последовательно активизирующихся генных единиц, и полиплоидия может рассматриваться как один из механизме создания такой системы. Исследования Маккей [ 58 ] показали, что эволюция пшеницы от примитивных ломкоколосых и спель-тоидных типов до прочноколосых и легко обмолачивающихся форм шла именно в направлении развития полигенной системы параллельно с возникновением полиплоидных видов. Кроме того, элементарные наборы хромосом, составляющие кариотип организма, при полиплоидизации или гибридизации не остаются тождественными, поскольку каждая хромосома с течением времени подвергается изменениям, что может и должно приводить к диплоидизации видов. При сохранении основного числа хромосом их генная структура изменяется. Мы хорошо знаем, что естественные полиплоидные формы и виды, прошедшие некоторый эволюционный путь развития, ведут себя функционально как диплоиды. Это последнее непосредственно сказывается на прохождении у них мейоза: как правило, образуются биваленты, а не мультиваленты, которые обычно характерны для только что полученных экспериментальных поли-лпидов. При этом вследствие увеличения числа хромосом при полиплоидии и генетической дифференциации между ранее гомологичными хромосомами будет повышаться индекс рекомбинации и изменчивости, что является источником видообразования. Кроме того, число жизнеспособных рекомбинантов при полиплоидизации возрастает, так как добавление, утеря или замена отдельных хромосом в мейозе происходит у полиплоидов с меньшим нарушением генетического баланса, чем у диплоидов. Такие особи могут быть исходными для образования производных видов. [23]
Отметим лишь следующее обстоятельство: в мелких хромосомах наблюдается в основном наличие гетерохроматиновых районов около центромеры. Это обнаружено у видов картофеля, помидор, капусты и ар. Данные особенности структуры хромосом, по всей видимости, не безразличны для их репродукции, образования нередуцированных гамет и полиплоидизации геномов. Однако посредством каких процессов это влияние осуществляется, пока не ясно. [24]
Строго говоря, все сказанное выше относится к оптимальным дозам аллелей. Но так как в организме существует довольно много ферментов, низкая активность которых лимитирует различные стадии роста, то одновременное увеличение активности всех лимитирующих ферментов приведет к увеличению гибридной мощности самого растения, а это возможно лишь при одновременной полиплоидизации всех генов. Растение будет иметь оптимальную плоидность ядра, если в нем можно создать такие соотношения аллелей по всем ферментам, лимитирующим рост организма, которые позволили бы наилучшим образом приблизиться к максимуму активности в каждом отдельном случае. [25]
Если дикие виды в условиях Новосибирска можно отнести к ранне - и среднеспелым формам, то посевной горох дополнен нами также позднеспелыми образцами с целью выяснения характера наследования этого признака на полиплоидном уровне. Такие лущильные зерновые сорта культурного гороха, как Торсдаг, Капитал, Урожайный, Виктория Мандорфская, Уладовский 387, Харьковский 131, Ранний майский и другие, включены нами в исследования не только с целью выяснения общей реакции на полиплоидизацию, но также для выяснения характера доминирования по таким признакам, как желтая окраска семядолей и округлость семян. С подобной же целью в опыты включены лущильные сорта Аляска, Ранний зеленый, Паули, Мергерт и др., но только имеющие уже один четкий рецессивный признак - зеленую окраску семядолей. Отличительной особенностью сортов Борец ГО-2040, Усатый 5, Победитель Г-33 является не только наличие у них 3 - 4 рецессивных признаков, которые на полиплоидном уровне можно использовать для гибридологического анализа, но также и то, что при скрещивании их с сортами типа Ранний майский, Аляска можно в ряде случаев получить высокий гетерозис по урожаю семян и вегетативных органов. Поэтому эти формы взяты в основном с целью последующей гибридизации на тетраплоидном уровне, которая, как нам кажется, может также представлять определенный интерес. [26]
При обсуждении роли полиплоидии в эволюции растений мы должны учитывать следующие обстоятельства. Полиплоиаизация хромосомного набора вида приводит не только к увеличению дозы соответствующего гена, что само по себе дает значительный фенотипический эффект и может иметь непосредственное адаптивное значение, но при дальнейших изменениях генов в результате мутагенеза полиплоидизация приводит к созданию новой генетической системы. Таким путем могут развиваться, вероятно, комплементарные и полигенные системы, имеющие определенное адаптивное значение. Известно, что каждый генокон - тролируемый процесс осуществляется системой, состоящей из многих генов. Продолжительность периода, в течение которого происходят метаболические процессы, также зависит от числа последовательно активизирующихся генных единиц, и полиплоидия может рассматриваться как один из механизме создания такой системы. Исследования Маккей [ 58 ] показали, что эволюция пшеницы от примитивных ломкоколосых и спель-тоидных типов до прочноколосых и легко обмолачивающихся форм шла именно в направлении развития полигенной системы параллельно с возникновением полиплоидных видов. Кроме того, элементарные наборы хромосом, составляющие кариотип организма, при полиплоидизации или гибридизации не остаются тождественными, поскольку каждая хромосома с течением времени подвергается изменениям, что может и должно приводить к диплоидизации видов. При сохранении основного числа хромосом их генная структура изменяется. Мы хорошо знаем, что естественные полиплоидные формы и виды, прошедшие некоторый эволюционный путь развития, ведут себя функционально как диплоиды. Это последнее непосредственно сказывается на прохождении у них мейоза: как правило, образуются биваленты, а не мультиваленты, которые обычно характерны для только что полученных экспериментальных поли-лпидов. При этом вследствие увеличения числа хромосом при полиплоидии и генетической дифференциации между ранее гомологичными хромосомами будет повышаться индекс рекомбинации и изменчивости, что является источником видообразования. Кроме того, число жизнеспособных рекомбинантов при полиплоидизации возрастает, так как добавление, утеря или замена отдельных хромосом в мейозе происходит у полиплоидов с меньшим нарушением генетического баланса, чем у диплоидов. Такие особи могут быть исходными для образования производных видов. [27]
Одно из эволюционных значений изменения числа хромосом, как известно, это образование изоляции между видами. При этом возникающая изоляция выявляется часто в почти полной нескрещиваемости видов с различными хромосомными числами. Для преодоления нескрещиваемости этих видов приходилось использовать предварительно полученные экспериментальные полиплоиды, именно тетраплоиды 24-хромосомных видов. При этом не только резко возрастала скрещиваемость, но и были возможны иные направления скрещиваний: не только с использованием дикого вида в качестве материнского растения, а культурного вида в качестве опылителя, что обычно приносило успех в селекционной практике при скрещивании с природными диплоидными видами, но и в обратном направлении, а именно с использованием в качестве материнского растения культурного вида, а в качестве опылителя - дикого вида, в его экспериментально полученной тетраплоидной форме. Однако они были в значительной степени стерильны при наличии сравнительно правильной конъюгации хромосом. Изоляция между вицами возникает из-за нарушений плодовитости у межвидовых гибридов, вследствие аномалий происхождения мейоза, отсутствия конъюгации хромосом или гибели пыльцы и зародышей в результате генетической несовместимости ядра и цитоплазмы и несбалансированности хромосом. Преодоление аномалий мейоза при межвидовой гибридизации и повышение плодовитости межвидовых гибридов, как сейчас хорошо известно, а впервые было экспериментально показано исследованиями Г.Д. Карпеченко [4-6], достигается полиплоидизацией набора хромосом, в силу чего возникают фертильные амфидиплоиды. В процессе эволюции часто имело место не только увеличение числа хромосом при полиплоидизации форм и становлении полиплоидных видов, но и гибридизация видов с одинаковыми или разными числами хромосом, сопутствуемая одновременно полиплоидизацией гибридного генома, наиболее часто за счет образования нередуцированных гамет. Частая встречаемость амфиплоидов в эволюции растений наглядно отразилась в различных таксонах растений возникновением новых вторичных чисел хромосом. В хорошо изученном за последние годы в этом отношении роде Clarkia ( сем. [28]
Одно из эволюционных значений изменения числа хромосом, как известно, это образование изоляции между видами. При этом возникающая изоляция выявляется часто в почти полной нескрещиваемости видов с различными хромосомными числами. Для преодоления нескрещиваемости этих видов приходилось использовать предварительно полученные экспериментальные полиплоиды, именно тетраплоиды 24-хромосомных видов. При этом не только резко возрастала скрещиваемость, но и были возможны иные направления скрещиваний: не только с использованием дикого вида в качестве материнского растения, а культурного вида в качестве опылителя, что обычно приносило успех в селекционной практике при скрещивании с природными диплоидными видами, но и в обратном направлении, а именно с использованием в качестве материнского растения культурного вида, а в качестве опылителя - дикого вида, в его экспериментально полученной тетраплоидной форме. Однако они были в значительной степени стерильны при наличии сравнительно правильной конъюгации хромосом. Изоляция между вицами возникает из-за нарушений плодовитости у межвидовых гибридов, вследствие аномалий происхождения мейоза, отсутствия конъюгации хромосом или гибели пыльцы и зародышей в результате генетической несовместимости ядра и цитоплазмы и несбалансированности хромосом. Преодоление аномалий мейоза при межвидовой гибридизации и повышение плодовитости межвидовых гибридов, как сейчас хорошо известно, а впервые было экспериментально показано исследованиями Г.Д. Карпеченко [4-6], достигается полиплоидизацией набора хромосом, в силу чего возникают фертильные амфидиплоиды. В процессе эволюции часто имело место не только увеличение числа хромосом при полиплоидизации форм и становлении полиплоидных видов, но и гибридизация видов с одинаковыми или разными числами хромосом, сопутствуемая одновременно полиплоидизацией гибридного генома, наиболее часто за счет образования нередуцированных гамет. Частая встречаемость амфиплоидов в эволюции растений наглядно отразилась в различных таксонах растений возникновением новых вторичных чисел хромосом. В хорошо изученном за последние годы в этом отношении роде Clarkia ( сем. [29]
Одно из эволюционных значений изменения числа хромосом, как известно, это образование изоляции между видами. При этом возникающая изоляция выявляется часто в почти полной нескрещиваемости видов с различными хромосомными числами. Для преодоления нескрещиваемости этих видов приходилось использовать предварительно полученные экспериментальные полиплоиды, именно тетраплоиды 24-хромосомных видов. При этом не только резко возрастала скрещиваемость, но и были возможны иные направления скрещиваний: не только с использованием дикого вида в качестве материнского растения, а культурного вида в качестве опылителя, что обычно приносило успех в селекционной практике при скрещивании с природными диплоидными видами, но и в обратном направлении, а именно с использованием в качестве материнского растения культурного вида, а в качестве опылителя - дикого вида, в его экспериментально полученной тетраплоидной форме. Однако они были в значительной степени стерильны при наличии сравнительно правильной конъюгации хромосом. Изоляция между вицами возникает из-за нарушений плодовитости у межвидовых гибридов, вследствие аномалий происхождения мейоза, отсутствия конъюгации хромосом или гибели пыльцы и зародышей в результате генетической несовместимости ядра и цитоплазмы и несбалансированности хромосом. Преодоление аномалий мейоза при межвидовой гибридизации и повышение плодовитости межвидовых гибридов, как сейчас хорошо известно, а впервые было экспериментально показано исследованиями Г.Д. Карпеченко [4-6], достигается полиплоидизацией набора хромосом, в силу чего возникают фертильные амфидиплоиды. В процессе эволюции часто имело место не только увеличение числа хромосом при полиплоидизации форм и становлении полиплоидных видов, но и гибридизация видов с одинаковыми или разными числами хромосом, сопутствуемая одновременно полиплоидизацией гибридного генома, наиболее часто за счет образования нередуцированных гамет. Частая встречаемость амфиплоидов в эволюции растений наглядно отразилась в различных таксонах растений возникновением новых вторичных чисел хромосом. В хорошо изученном за последние годы в этом отношении роде Clarkia ( сем. [30]
Страницы: 1 2 3