Половина - молекула
Cтраница 2
Получается, что одна половина молекулы является оптическим антиподом другой половины, и поэтому обе отклоняют плоскость поляризации на одинаковые углы, но в противоположных направлениях; в результате вся молекула в целом оптически неактивна. [16]
Проще говоря, каждая половина молекулы ДНК руководит синтезом своей второй половины и удерживает ее подле себя посредством водородных связей. Таким образом, из одной молекулы ДНК, которая разделилась на две одноцепочечные половины, образуются две двуспиральные молекулы, точно такие же, как и материнская молекула. Такой процесс удвоения, создания точной копии, или репликации, происходит со всеми молекулами ДНК, входящими в состав хромосом. В результате из одной хромосомы получаются две, которые являются копиями материнской хромосомы. Но иногда точность воспроизводства хромосом нарушается. Причиной этому могут стать либо взаимодействие С: ЯДер-ными; частицами, либо радиация, либо воздействие некоторых химических агентов, и тогда в процессе копирования ДНК происходят сбои, в результате чего дочерние хромосомы отличаются по какому-то фрагменту от материнской - происходит мутация. [17]
После насыщения кислородом одной половины молекулы гемоглобина возникает новое, внутреннее, напряженное состояние молекулы гемоглобина, которое вынуждает и вторую половину гемоглобина изменить конфор-мацию. Теперь еще две молекулы кислорода, по-видимому, по очереди связываются со второй половинкой молекулы гемоглобина, образуя оксигемоглобин. [18]
Формула показывает, что обе половины молекулы являются зеркальным отображением друг друга. [19]
Формула показывает, что обе половины молекулы являются зеркальным отображением друг друга. Если в левой половине молекулы гидроксил расположен у асимметрического атома углерода справа, а водород слева, то в правой половине молекулы, наоборот, гидроксил расположен слева, а водород справа. Обе эти половины молекулы мезовинной кислоты являются, таким образом, оптическими антиподами. Нетрудно понять поэтому причину оптической неактивности молекулы в целом. Легко понять также, почему мезовинная кислота не может быть разделена на свои оптические антиподы. Такого рода разделение было бы неизбежно связано с разрушением молекулы этой кислоты. [20]
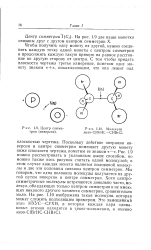 |
Центр симметрии ( инверсии.| Молекула лезо - С1ВгНС - СНВгС1. [21] |
Мы говорим, что одна половина молекулы получается из другой путем инверсии в центре симметрии. Хотя центро-симметричные молекулы встречаются довольно часто, молекул, обладающих только центром симметрии и не имеющих никакого другого элемента симметрии, сравнительно мало. На рис. 1.10 изображена такая молекула, которая может служить подходящим примером. [22]
 |
Влияние заместителей на сдвиг максимума поглощения в цианиновом красителе. [23] |
Разумеется, левая и правая половины молекулы здесь совершенно симметричны, положительный заряд рассредоточен между обоими азотами, а л-связи полиметиновой цепи делокализованы. [24]
 |
Влияние заместителей на сдвиг максимума поглощения в цианиновом красителе. [25] |
Разумеется, левая и правая половины молекулы здесь совершенно симметричны, положительный заряд рассредоточен между обоими азотами, а я-связи полиметиновой цепи делокализованы. [26]
Если представить теперь вращение двух половин молекулы вокруг линии связи С-С, то перекрывание р-орбит, которые лежат в одной плоскости и при этом образуют л-связь, уменьшается в той степени, в которой они выводятся из плоскости, и, наконец, при угле поворота около 90 перекрывание полностью исчезает - я-связь разрывается. Чтобы осуществить описанный поворот, необходимо, естественно, затратить энергию, освобождающуюся в результате перекрывания р-орбит. Если увеличить угол поворота еще больше ( сверх 90), то р-функции вновь начинают перекрываться и перекрывание достигает максимума ( минимум энергии) при повороте на 180 из исходного положения. [27]
Молекула витамина А соответствует точно половине молекулы Р - каротина, причем к месту разрыва по двойной связи присоединены ги-дроксильная группа и атом водорода. [28]
Молекула витамина А соответствует точно половине молекулы Р - каротина, причем к месту разрыва по двойной связи в центре молекулы присоединены гидроксильная группа и атом водорода; все двойные связи витамина А имеют транс-конфигурацию. [29]
Молекула витамина А соответствует точно половине молекулы р-каротина, причем к месту разрыва по двойной связи в центре молекулы присоединены гидроксильная группа и атом водорода; все двойные связи витамина А имеют транс-конформацию. [30]
Страницы: 1 2 3 4