Кодирующая последовательность
Cтраница 2
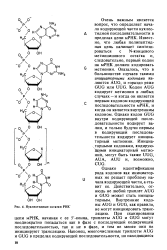 |
Нуклеотидные остатки РНК. [16] |
РНК, начиная с ее 5 -конца, триплеты AUG и GUG могут неоднократно попадаться как в фазе с последующей кодирующей последовательностью, так и не в фазе, и тем не менее они не инициируют трансляцию. [17]
Они показали, что во время трансляции РНК вируса энцефало-миокардита в определенном участке этой РНК существует трансляционный барьер, вызывающий значительную задержку во времени экспрессии последующей кодирующей последовательности. [18]
Надо сказать, что вне фазы ( рамки) считывания триплеты UAA, UAG и UGA в пределах кодирующей последовательности мРНК встречаются существенно чаще, чем в фазе считывания, где имеется, как правило, всего один терминирующий кодон на всю кодирующую последовательность. Поэтому обычно случайный сдвиг рамки в процессе элонгации не может привести к синтезу очень длинного неправильного полипептида и чаще всего приводит к скорой терминации этой неправильной трансляции. В некодирующих участках мРНК, включая межцистронные участки поли-цистронных РНК, частота терминирующих триплетов обычно также высока. [19]
Оперон - это группа сцепления структурных генов, наход щихся под контролем гена-оператора и гена-регулятора, а такя контролирующих элементов, узнаваемых продуктами регулято ] ного гена На рис 52 изображена схематичная структура твъ прокариотической клетки В его состав входят участки, располс женные перед кодирующей последовательностью ( лидерная 5 -о ( ласть) и после нее ( концевая З - область), синтез мРНК происходи в направлении от 5 -конца к З - концу Отсчет нуклеотидов в гена ведут от первого транскрибируемого нуклеотида, обозначаемог как 1 ( или 1), отсчет в противоположную сторону начинают - 1 Нд и РЪ на рисунке обозначают консервативные последова тельности Хогнесса и Прибноу соответственно Первая из ни представляет собою гексамер 5 ТТГАЦАЗ, локализующийся окол координаты - 35 Последовательность Хогнесса необходима дл. [20]
Геном FMDV представляет собой одноцепо-чечную РНК. Продукт кодирующей последовательности VPl-гена идентифицировали иммунологическими методами как часть слитого ( химерного) белка, синтез которого контролируется системой / ь-промотор - cl-penpeccop. Белок состоит из 396 аминокислотных остатков, содержит часть молекулы репликазы бактериофага MS2 и полноразмерный VPl-белок FMDV, благодаря чему он и индуцирует выработку нейтрализующих FMDV антител. [21]
Инициация в процессе биосинтеза белка означает не просто начало элонгации. Прежде всего, так как начало кодирующей последовательности мРНК не совпадает с началом полинуклеотидной цепи, а всегда находится, отступя от ее 5 -конца ( иногда на значительное расстояние), необходимо точное узнавание первого кодона на внутренней части цепи. Другими словами, именно инициация определяет фиксированную точку на матричном полинуклеотиде, с которой начинается отсчет триплетов без запятых ( см. гл. [22]
Инт-роны найдены в большинстве генов, кодирующих матричные РНК, и некоторых генах, кодирующих транспортные и рибосомные РНК. Во всех изученных случаях интроны копируются вместе с соседними кодирующими последовательностями как часть большого предшественника РНК. А затем интроны удаляются с помощью процесса, который называется сплайсингом. В результате интроны вы-щепляются, а информационные сегменты РНК сращиваются. Так возникает способная к функционированию молекула матричной РНК с непрерывной кодирующей последовательностью. Например, в гене человека, кодирующем глобин, существуют два интрона. Образовавшаяся после их удаления матричная РНК транспортируется из ядра в цитоплазму, где и происходит трансляция, т.е. считывание информации, необходимой для синтеза белка. [23]
Инт-роны найдены в большинстве генов, кодирующих матричные РНК, и некоторых генах, кодирующих транспортные и рибосомные РНК. Во всех изученных случаях интроны копируются вместе с соседними кодирующими последовательностями как часть большого предшественника РНК. А затем интроны удаляются с помощью процесса, который называется сплайсингом. В результате интроны вы-щепляются, а информационные сегменты РНК сращиваются. Так возникает способная к функционированию молекула матричной РНК с непрерывной кодирующей последовательностью. Например, в гене человека, кодирующем глобин, существуют два нитрона. Образовавшаяся после их удаления матричная РНК транспортируется из ядра в цитоплазму, где и происходит трансляция, т.е. считывание информации, необходимой для синтеза белка. [24]
Одна полинуклеотидная цепь мРНК не обязательно содержит только одну кодирующую последовательность. Для прокариотических мРНК очень обычно, что одна полинуклеотидная цепь включает кодирующие последовательности для нескольких белков. Такие мРНК получили название полицистронных мРНК ( происходит от термина цистрон, введенного С. Различные кодирующие последовательности ( цистроны) в пределах одной цепи мРНК обычно разделены некодирующими последовательностями. Такая внутренняя некодирующая последовательность начинается после терминаторного кодона предшествующего цистро-на; иногда в ее начале - имеется еще один терминаторный кодон, по-видимому, дублирующий терминаторный кодон цистрона на случай почему-либо не состоявшейся терминации. [25]
Позиционно-кандидатное картирование ( Positional-candidate gene cloning) Одна из стратегий идентификации гена заболевания, когда данные о продукте этого гена отсутствуют, но ген картирован в том же хромосомном районе, что и уже идентифицированные гены и EST. Сначала анализируют современные генетические и транскрипционные карты, с тем чтобы выявить кодирующие последовательности ( гены, внутригенные EST), находящиеся в этом районе, затем с помощью мутационного анализа или какими-либо другими методами выявляют экзоны, связанные с данным заболеванием. [26]
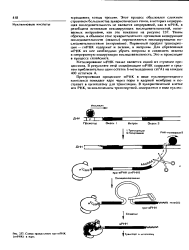 |
Схема процессннга пре-м РНК ( гяРНК в ядре. [27] |
Первичный продукт транскрипции - гяРНК содержит и экзоны, и нитроны. Для образования мРНК из нее необходимо убрать интроны и соединить экзоны в непрерывную кодирующую последовательность. Это и происходит в процессе сплайсинга. [28]
Следуя традиции, мы рассматриваем в этой главе в основном скользящее кодирование. В последнем параграфе изучаются погрешности рассмотренного ранее вида кодирования, возникающие при конечной длине кодирующей последовательности. [29]
Здесь мы получаем одно и то же решение, начиная с любого / - состояния XQ, которое достаточно похоже на ха. Следует отметить, что общий элемент / - состояний XQ и ха может быть практически невидимым из-за невероятно больших длин кодирующих последовательностей для XQ и ха. Например, XQ и ха могут иметь длину в 10000 знаков, причем только первые цифры этих последовательностей совпадают. [30]
Страницы: 1 2 3 4