Кодирующая последовательность
Cтраница 3
Процессинг первичных транскриптов SV40 включает те же реакции, что и процессинг большинства ядерных пре-м РНК. Все эти реакции в случае SV40 осуществляются клеточными ферментами и при помощи тех же механизмов, которые реализуются в незараженной клетке - Остановимся здесь лишь на сплайсинге, точнее, на его значении для образования индивидуальных мРНК в связи с особенностями организации кодирующих последовательностей в вирусном геноме. [31]
Процессинг первичных транскриптов SV40 включает те же реакции, что и процессинг большинства ядерных пре-м РНК эукариот: кэпирование 5 -конца, поли-аденилирование З - конца и сплайсинг. Все эти реакции в случае SV40 осуществляются клеточными ферментами и при помощи тех же механизмов, которые реализуются в незараженной клетке - Остановимся здесь лишь на сплайсинге, точнее, на его значении для образования индивидуальных мРНК в связи с особенностями организации кодирующих последовательностей в вирусном геноме. [32]
Этот белок закодирован участком РНК, начинающимся в конце С-цистрона, захватывающим всю 36-нуклеотидную вставку между С-цистроном и S-цистроном и заканчивающимся в пределах S-цистрона; рамка считывания этого перекрывающегося L-цистрона сдвинута вправо на один остаток ( 1 сдвиг), так что никакие его участки не транслируются при синтезе С-белка и S-белка. L-цистрон имеет свой инициаторный кодон AUG, не в фазе с кодо-нами С-цистрона, и, соответственно, свой терминаторный кодон UAA, не в фазе с кодонами S-цистрона. Использование перекрывающихся кодирующих последовательностей в пределах одной мРНК встречается, однако, не часто и свойственно, по-видимому, в основном вирусным системам, где экономия места для размещения цистронов играет особенно важную роль. [33]
Так как отрезки матричного полинуклеотида, непосредственно связанные с рибосомами, оказываются защищенными от действия нуклеаз, они могут быть выделены после нуклеазной обработки комплексов рибосома матрица. Длина таких отрезков была найдена равной от 20 до 60 нуклеотидных остатков. В то же время, длина кодирующей последовательности мРНК обычно превосходит 300 нуклеотидных остатков. Отсюда давно стало очевидно, что для считывания всей кодирующей последовательности мРНК рибосома должна последовательно пройти ( или, что то же самое, последовательно протащить через себя) матрицу, от 5 -концевой части кодирующей последовательности до ее 3 -концевой части. Другими словами, рибосома должна работать как лентопротяжный механизм. [34]
Сравнение аминокислотных последовательностей разных белковых токсинов показало, что белки некоторых штаммов имеют одинаковый домен, ответственный за токсичность. Кроме того, был субклонирован сегмент полной кодирующей последовательности, с которого синтезировался укороченный белок, в полной мере сохранивший свою токсичность. Таким образом, при последующих генноинженерных манипуляциях могут использоваться интактныи ген токсина, его фрагмент или химически синтезированный олигонуклеотид. [35]
В полицистроииых мРНК последовательности, кодирующие полипептидные цепи, разделены межцистронными нетранслируемыми участками ( спейсерами), длина к-рых варьирует в широких пределах. Между инициирующим ко-доном ближайшей к У-коицу мРНК транслируемой последовательности и этим концом обычно находится 50 - 200 нуклеотидных остатков. Конец мРНК отделен от терминирующего кодоиа ближайшей к нему кодирующей последовательности, как правило, значительно более длинным нетранслируемым участком ( до неск. Эти нетранслируемые области, по-видимому, необходимы для создания специфич. Наиб, хорошо изучена такая сигнальная последовательность у мРНК прокариот ( открыта Дж. Дальгарно и носит их имя), содержащая 3 - 7 нуклеотидиых остатков, расположенных перед инициирующим ко-доном ( т.е. ближе к 5 -коицу молекулы) каждой транслируемой последовательности. Последовательность Шайн-Дальгарно комплементарна З - концевому участку РНК малой субчастицы рибосом и, по всей вероятности, образует с ним в процессе инициации трансляции двухспиральный комплекс. [36]
Белок обладает биологической активностью лишь в том случае, если он имеет правильную трехмерную структуру. Синтез белков основан на информации, содержащейся в линейной, т.е. одномерной кодирующей последовательности ДНК. В соответствии с этой информацией рибосомы осуществляют сборку линейной, одномерной, последовательности аминокислот. Учитывая эти факты, объясните, каким образом в клетках могут формироваться биологически активные белки, обладающие специфической трехмерной структурой. Приведите какие-нибудь экспериментальные данные, подтверждающие ваши объяснения. [37]
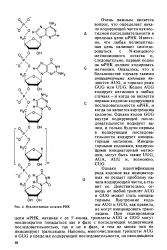 |
Нуклеотидные остатки РНК. [38] |
Известно, что любая полипептидная цепь начинает синтезироваться с N-концевого метионинового остатка и, следовательно, первый кодон на мРНК должен кодировать метионин. Оказалось, что в большинстве случаев такими инщиаторными кодонами являются AUG, и гораздо реже GUG или UUG. Кодон AUG кодирует метионин в любых случаях - и когда он является первым кодоном кодирующей последовательности мРНК, и когда он является внутренним кодоном. Однако кодон GUG внутри кодирующей последовательности кодирует ва-лин, и только будучи первым в кодирующей последовательности кодирует инициа-торный метионин. [39]
В осуществлении каждого из указанных процесов специфическое участие принимает ряд белков и нуклеиновых кислот, хотя конкретные молекулярные механизмы этих превращений еще не полностью раскрыты. Однако наибольший интерес исследователи проявляют к выяснению молекулярного механизма сплайсинга, который должен обеспечить, во-первых, постепенное и высокоточное вырезание нитронов из первичного транскрипта и, во-вторых, сшивание образующихся фрагментов-экзонов-конец в конец. Любые отклонения или смещения границ в процессе вырезания нитронов и сшивания экзонов даже на один нуклеотид могут привести не только к глубокому искажению смысла в кодирующих последовательностях, но и к нарушению передачи генетической информации и развитию патологии. [40]
Одна полинуклеотидная цепь мРНК не обязательно содержит только одну кодирующую последовательность. Для прокариотических мРНК очень обычно, что одна полинуклеотидная цепь включает кодирующие последовательности для нескольких белков. Такие мРНК получили название полицистронных мРНК ( происходит от термина цистрон, введенного С. Различные кодирующие последовательности ( цистроны) в пределах одной цепи мРНК обычно разделены некодирующими последовательностями. Такая внутренняя некодирующая последовательность начинается после терминаторного кодона предшествующего цистро-на; иногда в ее начале - имеется еще один терминаторный кодон, по-видимому, дублирующий терминаторный кодон цистрона на случай почему-либо не состоявшейся терминации. [41]
 |
Кодирующая последовательность ленцин-энкефалина с инициирующим и терминирующим кодонами. [42] |
Одним нз этапов синтеза искусственных двухцепочечных фрагментов ДНК является их клонирование в составе много копи иного вектора с тем, чтобы синтезированная последовательность сохранялась н при необходимости моглв быть наработана в достаточных количествах. С этой целью синтез двухцепочечных фрагментов осуществляют таким образом, чтобы онн содержали на концах участки, идентичные получающимся при расщеплении ДНК определенной рестрикционной эндонуклеазой. В квчестве примера на рисунке 209 приведена последовательность фрагмента ДНК, кодирующего короткий нейропептид - лейцин-энкефалнн. Наряду с собственно кодирующей последовательностью фрагмент содержит инициирующий АТС ( присоединенный к ней для обеспечения инициации трансляции) и терминирующий ТАА кодоны, в также выступающие одноцепочечные последовательности, соответствующие тем, которые образуются на концах ДНК при действии эндо-нуклеаз рестрикции EcoRl и BamHL. [43]
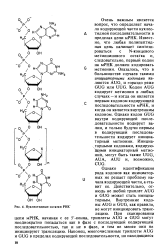 |
Нуклеотидные остатки РНК. [44] |
Известно, что любая полипептидная цепь начинает синтезироваться с N-концевого метионинового остатка и, следовательно, первый кодон на мРНК должен кодировать метионин. Оказалось, что в большинстве случаев такими инщиаторными кодонами являются AUG, и гораздо реже GUG или UUG. Кодон AUG кодирует метионин в любых случаях - и когда он является первым кодоном кодирующей последовательности мРНК, и когда он является внутренним кодоном. Однако кодон GUG внутри кодирующей последовательности кодирует ва-лин, и только будучи первым в кодирующей последовательности кодирует инициа-торный метионин. [45]
Страницы: 1 2 3 4