Сигнальная последовательность
Cтраница 3
Полиаденилирование З - конца первичного транскрипта включает ряд стадий и участие эндонуклеазы и полиаденилатполимеразы. Эндонуклеаза расщепляет мРНК вблизи специфической сигнальной последовательности ( 5) ААУААА ( 3), отличающейся высокой консервативностью. Полиаденилатполимераза синтезирует поли - А-конец ( от 20 до 250 нуклеотидов) начиная с точки распада. [31]
 |
Инсулярные гормоны. [32] |
На рибосомах шероховатого ретикулума р-клеток синтезируется препроинсулин, имеющий на W-конце молекулы гидрофобную последовательность из 16 аминокислот, с помощью которой белок проникает в эндоплазматический ретикулум. В просвете трубочек теряется сигнальная последовательность и образованный проинсулин транспортируется в аппарат Гольджи. Образуется инсулин, который накапливается в секреторных гранулах и секретируется при слиянии мембран гранул с плазматической мембраной клетки. [33]
Поскольку сигнальный пептид появляется от рибосомы, можно предполагать, что он прикреплен к микросомной мембране; это первый шаг в последовательности стадий, приводящих к переносу возникающей цепи через мембрану. В процессе или непосредственно после переноса сигнальная последовательность отделяется с помощью присоединенной к мембране протеазы. [34]
Для инициации транскрипции в нужном сайте необходим промотор, а для ее остановки - терминирующий кодон. Клонированный ген часто бывает лишен таких сигнальных последовательностей, и для его экспрессии в прокариотической клетке-хозяине нужно обеспечить и то, и другое. Кроме того, поскольку для решения большинства биотехнологических задач белок должен образовываться в больших количествах, необходимо использовать промотор, который позволял бы получить высокий уровень транскрипции ( сильный промотор) и распознавался РНК-полимеразой хозяйской клетки. Постоянная транскрипция клонированного гена истощает энергетические ресурсы хозяйской клетки, поэтому нужно использовать промоторы, работу которых можно регулировать либо с помощью специфических низкомолекулярных соединений, либо изменением температуры. [35]
Такие белки называют пробелками. После прохождения синтезированной полипептидной цепи через мембрану сигнальные последовательности отщепляются с помощью специальных связанных с мембранами протеаз. Кроме того, многие белки синтезируются в виде функционально - неактивных предшественников - пробелков. Это может иметь двоякое значение. Во-первых, это позволяет включить их функцию не сразу, а по достижении ими определенной мишени. Например, трипсин, который синтезируется поджелудочной железой в виде неактивного трипсиногена, активируется в кишечнике при действии специальной кишечной протеазы - энт. [36]
Если новый ген, содержащийся в рекомбинантной ДНК, обладает подходящими сигнальными последовательностями, указывающими начало и конец транскрипции, то такой ген будет транскрибироваться и транслироваться с образованием соответствующего белкового продукта. Многие гены из животных клеток уже были введены в бактерии, а бактериальные гены в свою очередь были встроены в эукариотиче-ские клетки. С помощью бактерий, в геномы которых введены соответствующие гены, можно получать применяемые в медицине белковые препараты-инсулин, гормон роста и интерфероны. [37]
Было сделано заключение, что трансляция первых кодонов мРНК приводит к синтезу N-концевой сигнальной последовательности, которая определяет присоединение рибосомы к мембране; в течение дальнейшей элонгации эта последовательность отщепляется на мембране, так что конечный продукт оказывается лишенным ее. Вслед за этим было показано, что такая N-концевая последовательность богата гидрофобными аминокислотными остатками; было естественно заключить, что гидрофобный характер сигнальной последовательности растущей полипептидной цепи способствует взаимодействию полипептида с мембраной. [38]
Несмотря на большое количество примеров, когда секреторные и внутримембранные белки синтезируются с отщепляемой сигнальной N-концевой последовательностью, это, по-видимому, не есть общее правило. Некоторые белки, как оказалось, тоже синтезируются на мембраносвязанных рибосомах, но без отщепления N-концевой или какой-либо иной сигнальной последовательности. К ним относится такой типичный секреторный белок, как овальбумин, а также мембранные белки цитохром Р-450, эпоксидгидратаза, ретинальный опсин, гликопротеид РЕа вируса Синдбис. В этих случаях растущая полипептидная цепь тоже, по-видимому, имеет сигнальную последовательность, индуцирующую присоединение транслирующей рибосомы к мембране эндоплазматического ретику-лума и ко-трансляционный транспорт белка в мембрану, но без сопутствующего процессинга. Для цитохрома Р-450 показано, что его N-концевая последовательность гидрофобна и напоминает сигнальную, но она сохраняется у зрелого белка. Однако N-концевые последовательности овальбумина и опсина не похожи на обычную сигнальную последовательность. Возможно, что либо сигнальную роль здесь выполняют специальные внутренние гидрофобные участки растущего полипептида, либо не столь гидрофобная N-концевая последовательность тоже в каких-то случаях может служить сигналом для присоединения к мембране. Как и в других случаях, взаимодействие с мембраной возможно лишь в течение трансляции; но не после нее; очевидно, сворачивание завершенной цепи как-то блокирует ( экранирует) сигнальную функцию соответствующего участка. [39]
 |
Освобождение белка из рибосомы. [40] |
Процесс закручивания полипептидной цепи происходит при помощи специальных белков - шапиронов ( гл. При синтезе мембранных и секреторных белков, начиная с Ж - конца полипептидной цепи, от 10 до 30 аминокислотных остатков образуют сигнальную последовательность, состоящую из гидрофобных аминокислот. В клетках существуют свободные и мембранно-свя-занные рибосомы, причем связывание их с мембраной ЭР определяется в основном сигнальной последовательностью растущего полипептида. В мембранах ЭР найдены два гликопротеина, получившие название рибофорины, которые специфически соединяются с сигнальной последовательностью полипептида. Это присоединение имеет более сложный характер. Оказалось, что в цитоплазме присутствуют специальные сигналузнающие структуры ( СУС), представляющие собой 1 IS рибонуклеопротеины. Они взаимодействуют с сигнальной последовательностью растущего полипептида, при этом элонгация временно прекращается. Синтезирующийся полипептид с СУС присоединяется к рибофоринам в мембране ЭР; при этом образуется мембранный канал, который иногда называют транслоконом. Элонгация возобновляется, но теперь она сопряжена с перемещением пептида через мембрану ЭР. После завершения синтеза полипептидной цепи под действием протеазы, которая носит название сигналаза, сигнальная последовательность отщепляется, а новосин-тезированный белок подвергается посттрансляционным модификациям или процессингу. Для большинства секреторных и мембранных белков процессинг сопряжен с транспортом через определенные компартменты. Так, гликозили-рование и ограниченный протеолиз начинаются уже в ЭР и продолжаются в аппарате Гольджи. Этот компартмент состоит из 12 - 15 тарелок, сложенных в стопку. Сторона, ориентированная на ЭР, называется цис-стороной, а в направлении цитоплазматической мембраны - / и / анс-стороной. Новосинтези-рованные белки поступают на цис-сторону аппарата Гольджи и перемещаются на его транс - сторону, пересекая все тарелки, причем по мере движения происходит их химическая модификация. Эта модификация имеет огромное значение, так как она, в частности, определяет следование новосинтезированного белка к месту функционирования. [41]
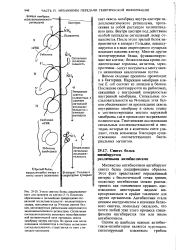
Весьма сходные процессы происходят и в бактериях. Последние синтезируются на рибосомах, связанных с внутренней поверхностью внутренней мембраны. Сигнальные последовательности на N-концах этих белков обеспечивают их прохождение сквозь внутреннюю мембрану и сквозь стенку к соответствующим местам наружной мембраны, где и происходит встраивание полипептида. Генетические исследования сигнальных последовательностей и пеп-тидаз, которые их в конечном счете удаляют, стали возможны благодаря существованию соответствующих бактериальных мутантов. [43]
Следует подчеркнуть, что остановка элонгации по выходе сигнальной последовательности из рибосомы представляется очень важной для нормальной секреции ( или вхождения4 мембрану) синтезируемого белка. Следовательно, остановка элонгации обеспечивает необходимое ожидание сигнальной последовательности в нужной экспонированной конформации для причаливания к мембране. [44]
Кроме секретируемых белков, растущие полипептидные цепи ряда встроенных в мембрану белков также характеризуются временной сигнальной N-концевой последовательностью. Одним из первых изученных примеров такого рода был гликопротеид вируса везикулярного стоматита, который вместе с хозяйской мембраной участвует в построении вирусной оболочки. Этот белок, как оказалось, синтезируется с N-концевой сигнальной последовательностью, очень похожей на таковую секретируемых пребелков; сигнальная последовательность необходима для присоединения транслирующей рибосомы к мембране эндоплазматического ретикулума; дальнейший синтез белка идет, таким образом, на мембраносвязанных рибосомах; в ходе элонгации N-концевая последовательность из 16 аминокислотных остатков отщепляется в мембране. Другими словами, все это не отличимо от ситуации в случае водорастворимых секретируемых белков. [45]
Страницы: 1 2 3 4