Сигнальная последовательность
Cтраница 4

После того, как полипептидные цепи отделились от рибосомы, они могут тотчас же приобретать свойственную им вторичную, третичную или четвертичную структуру ( разд. Если рибосома прикреплена к ЭР, то белок поступает в ЭР, откуда затем траспорти-руется к месту назначения. Первая часть растущей полипептидной цепи состоит при этом из сигнальной последовательности аминокислот, которая соответствует определенному рецептору в мембране ЭР, что обеспечивает присоединение рибосомы к ЭР. После того как белок оказывается внутри ЭР, сигнальная последовательность отделяется от него, и белковая молекула свертывается, приобретая свою окончательную форму. [47]
Для клонирования эукариотических структурных генов необходимы специальные методики. Прокариоты не способны удалять интроны из первичных РНК-транскриптов, поэтому правильная трансляция эукариотических мРНК в бактериальной клетке невозможна. Более того, экспрессия эукариотической ДНК может осуществляться только при наличии прокариотиче-ских сигнальных последовательностей, регулирующих транскрипцию и трансляцию. Концевые участки эукариотических мРНК особым образом модифицированы: их 5 -концы кэ-пированы ( содержат кэп из остатка G, часто метилированного), а 3 -концы полиаденилиро-ваны ( содержат ро. [48]
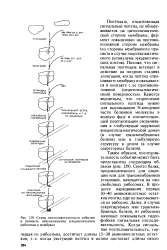 |
Схема последовательности событий в процессе присоединения эукариотической рибосомы к мембране. [49] |
Синтез белка, предназначенного для секреции или для трансмембранной установки, начинается на сво-свободных рибосомах. В процессе наращивания первых 30 - 40 аминокислотных остатков пептид еще не высовывается из рибосом. Далее, в случае секретируемых или трансмембранных белков, из рибосомы начинает показываться гидрофобная сигнальная последовательность. [50]
Процесс закручивания полипептидной цепи происходит при помощи специальных белков - шапиронов ( гл. При синтезе мембранных и секреторных белков, начиная с Ж - конца полипептидной цепи, от 10 до 30 аминокислотных остатков образуют сигнальную последовательность, состоящую из гидрофобных аминокислот. В клетках существуют свободные и мембранно-свя-занные рибосомы, причем связывание их с мембраной ЭР определяется в основном сигнальной последовательностью растущего полипептида. В мембранах ЭР найдены два гликопротеина, получившие название рибофорины, которые специфически соединяются с сигнальной последовательностью полипептида. Это присоединение имеет более сложный характер. Оказалось, что в цитоплазме присутствуют специальные сигналузнающие структуры ( СУС), представляющие собой 1 IS рибонуклеопротеины. Они взаимодействуют с сигнальной последовательностью растущего полипептида, при этом элонгация временно прекращается. Синтезирующийся полипептид с СУС присоединяется к рибофоринам в мембране ЭР; при этом образуется мембранный канал, который иногда называют транслоконом. Элонгация возобновляется, но теперь она сопряжена с перемещением пептида через мембрану ЭР. После завершения синтеза полипептидной цепи под действием протеазы, которая носит название сигналаза, сигнальная последовательность отщепляется, а новосин-тезированный белок подвергается посттрансляционным модификациям или процессингу. Для большинства секреторных и мембранных белков процессинг сопряжен с транспортом через определенные компартменты. Так, гликозили-рование и ограниченный протеолиз начинаются уже в ЭР и продолжаются в аппарате Гольджи. Этот компартмент состоит из 12 - 15 тарелок, сложенных в стопку. Сторона, ориентированная на ЭР, называется цис-стороной, а в направлении цитоплазматической мембраны - / и / анс-стороной. Новосинтези-рованные белки поступают на цис-сторону аппарата Гольджи и перемещаются на его транс - сторону, пересекая все тарелки, причем по мере движения происходит их химическая модификация. Эта модификация имеет огромное значение, так как она, в частности, определяет следование новосинтезированного белка к месту функционирования. [51]
Кроме секретируемых белков, растущие полипептидные цепи ряда встроенных в мембрану белков также характеризуются временной сигнальной N-концевой последовательностью. Одним из первых изученных примеров такого рода был гликопротеид вируса везикулярного стоматита, который вместе с хозяйской мембраной участвует в построении вирусной оболочки. Этот белок, как оказалось, синтезируется с N-концевой сигнальной последовательностью, очень похожей на таковую секретируемых пребелков; сигнальная последовательность необходима для присоединения транслирующей рибосомы к мембране эндоплазматического ретикулума; дальнейший синтез белка идет, таким образом, на мембраносвязанных рибосомах; в ходе элонгации N-концевая последовательность из 16 аминокислотных остатков отщепляется в мембране. Другими словами, все это не отличимо от ситуации в случае водорастворимых секретируемых белков. [52]
Было сделано заключение, что трансляция первых кодонов мРНК приводит к синтезу N-концевой сигнальной последовательности, которая определяет присоединение рибосомы к мембране; в течение дальнейшей элонгации эта последовательность отщепляется на мембране, так что конечный продукт оказывается лишенным ее. Вслед за этим было показано, что такая N-концевая последовательность богата гидрофобными аминокислотными остатками; было естественно заключить, что гидрофобный характер сигнальной последовательности растущей полипептидной цепи способствует взаимодействию полипептида с мембраной. [53]
Гликозилированию секретпруемых белков предшествует протеолитнч. Наиб, распространенные сигнальные последовательности включают 23 аминокислотных остатка. Характерные особенности этих последовательностей - наличие на конце короткого положительно заряженного участка, за к-рым следует гидрофобный участок, содержащий от 7 до 14 аминокислотных остатков. Сигнальные последовательности завершаются консервативным по длине ( 5 - 7 остатков) гидрофильным участком, на С-конце к-рого чаще всего находятся остатки аланина, глицина, серина, треонина, цистеина или глутамина. [54]
ПОСТ-ТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ белков, катализируемые специфич. РНК или после нее; участвует в формировании пространств, структуры белков и их функц. N-конце-вых сигнальных последовательностей, обеспечивающих перенос секреторных белков через мембрану; гликози-лирование - последоват. [55]
ПОСТ-ТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ белков, катализируемые специфич. РНК или после нее; участвует в формировании пространств. N-конце-вых сигнальных последовательностей, обеспечивающих перенос секреторных белков через мембрану; тликози-лирование - последоват. [56]
Восстановлением дисульфидных связей и их последующим реокислением был получен проинсулин с - 70 % - ным выходом. На рис. 2 - 41 приведена первичная структура проинсулина свиньи, состоящего из 84 аминокислот. В процессе биосинтеза на первом этапе образуется фрагмент, содержащий в соответствии с так называемой сигнальной гипотезой ( разд. N-концевую сигнальную последовательность с информацией о том, что последующая последовательность проиисулина строится прямо на мембране эндоплазматического ретикулума и что этот инсулиновый предшественник уже в процессе синтеза должен уходить через мембрану по внутренним каналам ЭР. Еще во время биосинтеза сигнальный пептид отщепляется особой сигнальной пептидазой. Проинсулин затем идет в аппарат Гольджи, где расщепляется на инсулин и С-пелтид и в кристаллической форме в присутствии цинка накапливается в визикулах. При соответствующем физиологическом раздражении происходит экзоцитоз и содержимое визикул попадает в кровь. [57]
Весьма сходные процессы происходят и в бактериях. Последние синтезируются на рибосомах, связанных с внутренней поверхностью внутренней мембраны. Сигнальные последовательности на N-концах этих белков обеспечивают их прохождение сквозь внутреннюю мембрану и сквозь стенку к соответствующим местам наружной мембраны, где и происходит встраивание полипептида. Генетические исследования сигнальных последовательностей и пеп-тидаз, которые их в конечном счете удаляют, стали возможны благодаря существованию соответствующих бактериальных мутантов. [58]
Страницы: 1 2 3 4