Постсинаптический потенциал
Cтраница 4
Уменьшение скорости и амплитуды нервных импульсов, когда они достигают окончаний аксонов, возникает из-за заметного уменьшения диаметра окончаний волокон. Следовательно, пресинаптических потенциалов самих по себе еще далеко не достаточно, чтобы вызвать постсинаптический потенциал. [46]
Прикосновение к сифону ведет к возбуждению группы сенсорных нейронов. Эти нейроны образуют возбуждающие синапсы на других нейронах, которые непосредственно управляют мышцами, втягивающими жабру. Реакцию последней группы нейронов на импульсы от сенсорных нейронов можно регистрировать внутриклеточным электродом; оказывается, во время привыкания величина постсинаптического потенциала при повторном возбуждении уменьшается. При сенситизации наблюдается обратный эффект - постсинаптический потенциал возрастает. [47]
Со времени открытия более полувека тому назад аце-тилхолина арсенал трансмиттеров пополнился примерно десятком других органических соединений, выступающих в качестве синаптических веществ-передатчиков. Они накапливаются в синаптических пузырьках, скользят к пост-синаптической мембране на входе нервной клетки, где соединяются с протеиновыми рецепторами и на короткое время селективно изменяют проводимость мембраны для определенных ионов. Длящиеся от 10 до 20 миллисекунд постсинаптические де - и гиперполяризации в несколько милливольт мембранной поляризации ( внутриклеточная - около 80 милливольт по отношению к внеклеточной) суммируются по времени и пространству и ведут к дальнейшему возбуждению в форме акциональных потенциалов, если не будет повышен критический порог поляризации ( например, в 65 милливольт) у аксонного выступа. Если темпоральная суммация локальных возбуждающих постсинаптических потенциалов ( ВПСП) и тормозящих постсинаптических потенциалов ( ТПСП) является следствием сукцессивных ( наследственных) синаптических возбуждений, то локальная суммация зависит от одновременного поступления потенциалов через различные контактирующие участки. [48]
Медиатор быстро диффундирует через щель, воздействуя на мембрану иннервируемого мышечного волокна. Та часть мембраны этого волокна, которая непосредственно граничит с нервным окончанием, называется постринаптиче-ской. От мембраны, покрывающей остальную часть мышечного волокна, постсинапти-ческая мембрана отличается тем, что не содержит электрически возбудимых ионных каналов и потому не способна к генерации потенциала действия. По своей природе он аналогичен возбуждающим постсинаптическим потенциалам, возникающим при передаче возбуждения с одной нервной клетки на другую. ПКП порождает генерацию потенциала действия в мышечном волокне. Наличие химического звена в механизме нервно-мышечной передачи делает понятным два общих свойства синапсов: 1) возбуждение проводится через синапс только в одном направлении - с нерва на мышцу; 2) возбуждение проводится через синапс значительно медленнее, чем по нервному волокну. [49]
Прикосновение к сифону ведет к возбуждению группы сенсорных нейронов. Эти нейроны образуют возбуждающие синапсы на других нейронах, которые непосредственно управляют мышцами, втягивающими жабру. Реакцию последней группы нейронов на импульсы от сенсорных нейронов можно регистрировать внутриклеточным электродом; оказывается, во время привыкания величина постсинаптического потенциала при повторном возбуждении уменьшается. При сенситизации наблюдается обратный эффект - постсинаптический потенциал возрастает. [50]
Со времени открытия более полувека тому назад аце-тилхолина арсенал трансмиттеров пополнился примерно десятком других органических соединений, выступающих в качестве синаптических веществ-передатчиков. Они накапливаются в синаптических пузырьках, скользят к пост-синаптической мембране на входе нервной клетки, где соединяются с протеиновыми рецепторами и на короткое время селективно изменяют проводимость мембраны для определенных ионов. Длящиеся от 10 до 20 миллисекунд постсинаптические де - и гиперполяризации в несколько милливольт мембранной поляризации ( внутриклеточная - около 80 милливольт по отношению к внеклеточной) суммируются по времени и пространству и ведут к дальнейшему возбуждению в форме акциональных потенциалов, если не будет повышен критический порог поляризации ( например, в 65 милливольт) у аксонного выступа. Если темпоральная суммация локальных возбуждающих постсинаптических потенциалов ( ВПСП) и тормозящих постсинаптических потенциалов ( ТПСП) является следствием сукцессивных ( наследственных) синаптических возбуждений, то локальная суммация зависит от одновременного поступления потенциалов через различные контактирующие участки. [51]

Электронно-микроскопические исследования показали, что пресинаптические окончания всегда содержат синаптические пузырьки или везикулы, каждая из которых содержит один квант медиатора. Действительно, имеются убедительные биохимические данные, что вещества, рассматриваемые в качестве химических медиаторов, содержатся в синаптических пузырьках. Более того, расчеты количества медиатора, содержащегося в одном пузырьке, и количество молекул медиатора, необходимых для создания постсинаптического эффекта, аналогичного действию одного кванта, совпадают. Таким образом, совокупность имеющихся данных свидетельствует о том, что как спонтанные миниатюрные постсинаптические потенциалы, так и постсинаптические потенциалы обусловлены выходом в синаптическую щель медиатора, содержащегося в синаптических пузырьках. Этот процесс ( экзоцитоз) заключается в том, что пузырек, подойдя к внутренней поверхности мембраны пресинаптического окончания при наличии Са 2, сливается с пресинаптической мембраной. В результате происходит опорожнение пузырька в синаптическую щель. После спадения пузырька окружающая его мембрана включается в мембрану пресинаптического окончания, увеличивая его поверхность. В дальнейшем ( в результате процесса эндоцитоза) небольшие участки пресинаптической мембраны впячиваются внутрь, вновь образуя пузырьки, которые впоследствии снова способны включать медиатор и вступать в цикл его высвобождения. [53]
Существенную роль играет изменение ( переключение) связей между элементами сетчатки. Как установлено, в темноте площадь возбудительного центра рецептивного поля ганглиозной клетки увеличивается вследствие ослабления или снятия кольцевого торможения. При этом к одной биполярной клетке подключено большое число фоторецепторов и большее их число конвергирует на ганглиозную клетку. Вследствие этого при очень слабом фоновом освещении и в темноте вследствие пространственной суммации возбуждающий постсинаптический потенциал увеличивается, а порог реакций на свет ганглиозных клеток снижается. [54]
По нашим данным, предварительное ( за 2 - 3 ч) введение кошкам больших доз гепарина ( 50 мг / кг) предотвращало угнетение первичных ответов коры при прямой аппликации яда кобры ( 1 %) на поверхность соматосенсорной зоны. Механизм защиты гепарином именно деполяризующих синапсов пока не ясен. Я - МихельсО Н, Э. В. Зеймаль, 1970), что может обусловить относительную устойчивость холинорецептивной мембраны к действию яда. Очевидно, в условиях защиты гепарином деполяризующих синапсов яд в меньшей степени угнетает возбуждающий постсинаптический потенциал апикальных дендритов, что проявляется в устойчивости отрицательного колебания первичных ответов к действию яда кобры. Другой возможный механизм защитного действия гепарина может быть связан с его хорошо выраженными комплексообразующими свойствами ( А. А. Маркосян, 1969), в результате чего инактивация яда происходит путем связывания его гепарином. [55]
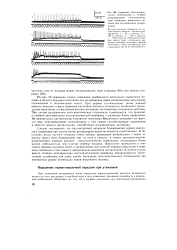 |
Суммация постсинапти-ческих потенциалов и стойкая деполяризация постсинаптиче-ской мембраны мышечного волокна при пессимальном раздражении. [56] |
Следует подчеркнуть, что рассмотренный механизм блокирования нервно-мышечного соединения при частом ритме раздражения нерва не является единственным. В тех случаях, когда частота стимулов очень высока, проведение возбуждения с нерва на мышцу может быть блокировано еще на пути к синапсу, в тонких пресинаптических разветвлениях нервных волокон - пресинаптических терминалях, обладающих более низкой лабильностью, чем толстые нервные волокна. Нарушение проведения в этих тонких нервных волокнах ведет к прекращению поступления нервных импульсов к нервному окончанию и тем самым к прекращению выделения ацетилхолина. В данном случае вместо стойкой деполяризации постсинаптической мембраны обнаруживается значительное ослабление или даже полное выпадение постсинаптических потенциалов при неизменном уровне потенциала покоя мышечного волокна. [57]
Электронно-микроскопические исследования показали, что пресинаптические окончания всегда содержат синаптические пузырьки или везикулы, каждая из которых содержит один квант медиатора. Действительно, имеются убедительные биохимические данные, что вещества, рассматриваемые в качестве химических медиаторов, содержатся в синаптических пузырьках. Более того, расчеты количества медиатора, содержащегося в одном пузырьке, и количество молекул медиатора, необходимых для создания постсинаптического эффекта, аналогичного действию одного кванта, совпадают. Таким образом, совокупность имеющихся данных свидетельствует о том, что как спонтанные миниатюрные постсинаптические потенциалы, так и постсинаптические потенциалы обусловлены выходом в синаптическую щель медиатора, содержащегося в синаптических пузырьках. Этот процесс ( экзоцитоз) заключается в том, что пузырек, подойдя к внутренней поверхности мембраны пресинаптического окончания при наличии Са 2, сливается с пресинаптической мембраной. В результате происходит опорожнение пузырька в синаптическую щель. После спадения пузырька окружающая его мембрана включается в мембрану пресинаптического окончания, увеличивая его поверхность. В дальнейшем ( в результате процесса эндоцитоза) небольшие участки пресинаптической мембраны впячиваются внутрь, вновь образуя пузырьки, которые впоследствии снова способны включать медиатор и вступать в цикл его высвобождения. [58]
Нервные сигналы переходят от клетки к клетке через синапсы, которые могут быть электрическими ( щелевые контакты) или химическими. В химическом синапсе деполяризация пресинаптической мембраны в результате прибытия нервного импульса открывает потенциал-зависимые кальциевые каналы, вызывая тем самым приток Са2 в клетку, что приводит к освобождению нейромедиатора из синоптических пузырьков. Медиатор диффундирует в синоптическую щель и связывается с рецептурными белками в мембране постсинаптической клетки; в конечном итоге медиатор удаляется из синоптической щели путем диффузии, ферментативного расщепления или обратного поглощения выделившей его клеткой. Через рецепторные белки, образующие лиганд-зависимые каналы, реализуется быстрый постсинаптический эффект нейромедиатора-открытие каналов приводит к возникновению возбуждающего или тормозного постсинаптического потенциала в соответствии с ионной специфичностью каналов. При участии рецепторов, сопряженных с ферментами, например с аденилатциклазой, обычно осуществляются медленные и более продолжительные эффекты. [59]
 |
Пространственная и временная суммация. [60] |
Страницы: 1 2 3 4 5