Рекомбинационный процесс
Cтраница 1
Рекомбинационные процессы играют также ведущую роль в эволюции строения геномов в целом. Дело в том, что перестройки генетического материала часто можно объяснить рекомбинацией между гомологичными последовательностями, оказавшимися в негомологичном положении ( роль таких последовательностей могут выполнять, например, мобильные генетические элементы; см. гл. В результате этого процесса генетический материал одной из гомологичных хромосом делегирует, но в другой хромосоме возникает дупликация. Считается, что такие дупликации играют важную роль в возникновении родственных, но различных генов, поскольку присутствие в геноме лишних копий какого-либо гена позволяет им сравнительно свободно изменяться, что, в принципе, может привести к возникновению новых функций белка - продукта гена. [1]
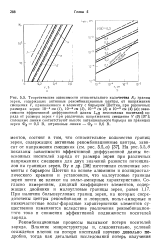
Рекомбинационные процессы вызывают потери носителей заряда. [3]
Рекомбинационные процессы в арсениде галлия, фосфиде галлия и карбиде кремния, имеющих большую ширину запрещенной зоны, сопровождаются выделением энергии в виде квантов света, которые частично поглощаются объемом полупроводника, а частично излучаются в окружающее пространство. Поэтому внешний квантовый выход, фиксируемый зрительно, всегда меньше внутреннего. [4]

Рекомбинационные процессы, в которых участвуют сегменты ДНК, не обнаруживающие заметной генетической гомологии, называют негомологичной рекомбинацией. Так же как и сайт-специфическая рекомбинация, она представляет собой интеграционную форму рекомбинации, т.е. не обмен, а соединение ДНК. Негомологичная рекомбинация независима от гена гее А. Молекулярный механизм негомологичной рекомбинации еще не вполне выяснен. [6]
Рекомбинационные процессы играют также ведущую роль в эволюции строения геномов в целом. Дело в том, что перестройки генетического материала часто можно объяснить рекомбинацией между гомологичными последовательностями, оказавшимися в негомологичном положении ( роль таких последовательностей могут выполнять, например, мобильные генетические элементы; см. гл. В результате этого процесса генетический материал одной из гомологичных хромосом делети-рует, но в другой хромосоме возникает дупликация. Считается, что такие дупликации играют важную роль в возникновении родственных, но различных генов, поскольку присутствие в геноме лишних копий какого-либо гена позволяет им сравнительно свободно изменяться, что, в принципе, может привести к возникновению новых функций белка - продукта гена. [7]
Все рекомбинационные процессы относятся к рекомбинации электронов и ионов аргона и представлены реакциями (13.43), (13.48) и (13.49), идущими влево. [8]
Анализ рекомбинационных процессов показал, что с увеличением концентрации глубоких локальных центров время жизни носителей заряда уменьшается. В кристалле же всегда есть область, в которой концентрация локальных центров велика. Этой областью является поверхность кристалла. В реальных условиях, поскольку полупроводник контактирует с внешней средой, на его поверхности всегда присутствуют слои окисла и адсорбированные примеси. Поверхностные состояния бывают двух типов: быстрые и медленные. Быстрые состояния характерны временами захвата на них электронов и дырок порядка нескольких микросекунд, а для медленных состояний эти времена составляют обычно миллисекунды, а в ряде случаев они достигают даже нескольких часов. [9]
Особенности рекомбинационных процессов являются сложными и, многообразными и во многом определяют характеристики и параметры фотоприемников. [10]
При рекомбинационных процессах иногда наблюдаются свечения с длительностью 10 - 7 - - 10 - 5 сек. [11]
Какой из рекомбинационных процессов будет доминировать, зависит в основном от концентраций Rx и R2, устанавливающихся в ходе реакции. Концентрации эти в свою очередь определяются скоростями, с которыми реагируют радикалы Rx и R2 в реакциях продолжения цепи. [12]
Какой из рекомбинационных процессов будет доминировать, зависит в основном от концентраций Rx и R2, устанавливающихся в ходе реакции. Концентрации эти в свою очередь определяются скоростями, с которыми реагируют радикалы RJ и R2 в реакциях продолжения цепи. [13]
За счет рекомбинационных процессов он излучает свет. Коллектор включают в обратном направлении. Излучение с эмиттерного перехода поглощается в области коллекторного перехода, в результате в коллекторе и во внешней цепи проходит ток. Для работы оптотранзистора необходимо, чтобы в базе не поглощалось излучение, испускаемое эмиттерным p - n - переходом. Для электрической изоляции входной и выходной цепей в базе параллельно с p - n - переходами ( рис. 1.56) создают высокоомный слой. [14]
Медленное протекание рекомбинационных процессов позволяет пренебречь ими на периферии возбужденной области в течение сотен микросекунд. При этом ионная проводимость остается постоянной и уступает примерно на порядок максимальному значению т ах. [15]
Страницы: 1 2 3 4